





























I. Tổng quan
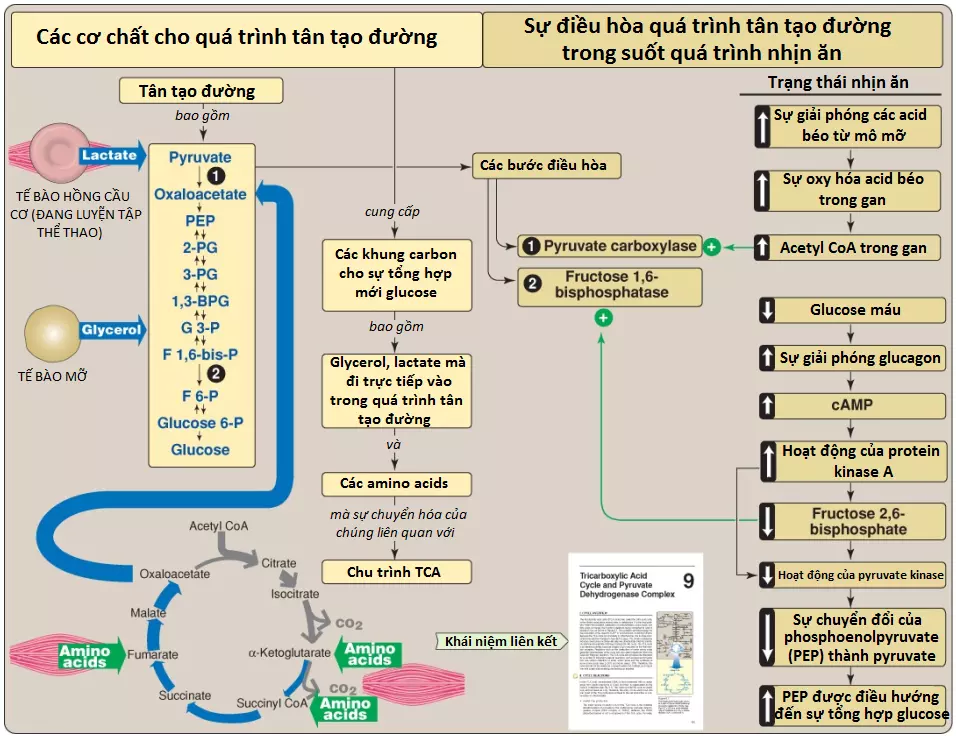
Một số mô như não, hồng cầu, tủy thận, thủy tinh thể và võng mạc của mắt, các tinh hoàn và cơ xương đang luyện tập thể thao, cần một sự cung cấp glucose liên tục như là nguyên liệu cho chuyển hóa. Glycogen gan, một nguồn glucose thiết yếu sau bữa ăn, có thể đáp ứng các nhu cầu này trong vòng dưới 24 giờ khi không có carbohydrate nạp vào trong chế độ ăn. Tuy nhiên, trong suốt quá trình nhịn ăn kéo dài, các tích trữ glycogen gan bị cạn kiệt và glucose sẽ được tạo thành từ các tiền thân không phải là carbohydrate. Sự hình thành glucose không xảy ra bởi một sự đảo ngược đơn giản của quá trình đường phân bởi vì cân bằng tổng quan của quá trình đường phân ưu tiên một cách mạnh mẽ sự hình thành pyruvate. Thay vào đó, glucose được tổng hợp mới bởi một con đường chuyên biệt được gọi là quá trình tân tạo đường (gluconeogenesis), quá trình này yêu cầu cả các enzyme ty thể và bào tương. Các sự suy giảm các enzymes tân tạo đường sẽ gây ra sự hạ đường huyết. Trong suốt một quá trình nhịn ăn qua đêm, khoảng 90% sự tân tạo đường xảy ra trong gan với khoảng 10% còn lại xảy ra trong các thận. Tuy nhiên, trong suốt quá trình nhịn ăn kéo dài khoảng 48 giờ hoặc lâu hơn thì các thận trở thành các cơ quan sản xuất glucose chính, đóng góp khoảng 40% tổng sự sản xuất glucose. Ruột non cũng có thể tạo ra glucose. Hình 1 cho thấy mối liên hệ của quá trình tân tạo đường với các con đường chuyển hóa năng lượng thiết yếu khác.

II. Các cơ chất
Các tiền thân tân tạo đường (gluconeogenic precursors) là các phân tử mà có thể được sử dụng để tạo ra một sự tổng hợp glucose toàn phần. Các tiền thân tân tạo đường quan trọng nhất là glycerol, lactate và các α-keto acids thu được từ sự chuyển hóa của các amino acids tạo đường (tạo glucose). Tất cả trừ hai amino acids (leucine và lysine) là các amino acids tạo đường.
A. Glycerol
Glycerol được giải phóng trong suốt quá trình thủy phân của triacylglycerols (TAGs) trong mô mỡ và được vận chuyển bởi máu đến gan. Glycerol được phosphoryl hóa bởi glycerol kinase thành glycerol 3-phosphate, thành phần mà bị oxy hóa bởi glycerol 3-phosphate dehydrogenase thành dihydroxyacetone phosphate, một trung gian của quá trình đường phân (glycolysis) và quá trình tân tạo đường (gluconeogenesis).
B. Lactate
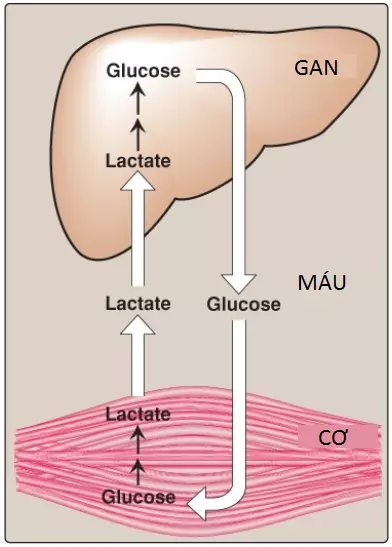
Lactate từ quá trình đường phân kỵ khí (anaerobic glycolysis) được giải phóng vào trong máu bởi cơ xương đang luyện tập thể thao và bởi các tế bào hồng cầu, các tế bào mà thiếu ty thể. Trong chu trình Cori, lactate này được hấp thu bởi gan và được oxy hóa thành pyruvate, thành phần mà được chuyển thành glucose, sau đó được giải phóng lại vào trong hệ thống tuần hoàn (Hình 2).
C. Các amino acids
Các amino acids được sản xuất bởi quá trình thủy phân các protein mô là các nguồn glucose chính trong suốt quá trình nhịn ăn. Sự chuyển hóa của chúng tạo ra các α-keto acids như pyruvate mà được chuyển thành glucose hay α-ketoglutarate mà có thể đi vào trong chu trình tricarboxylic acid (tricarboxylic acid cycle) (chu trình TCA [TCA cycle]) và hình thành nên oxaloacetate (OAA), một tiền thân trực tiếp của phosphoenolpyruvate (PEP). (Chú ý: Acetyl coenzyme A [CoA] và các hợp chất mà chỉ có thể chuyển thành acetyl CoA [như acetoacetate, lysine và leucine], không thể tạo ra sự tổng hợp glucose toàn phần. Điều này là bởi vì bản chất không thể đảo ngược của phức hợp pyruvate dehydrogenase [pyruvate dehydrogenase complex – PDHC], thành phần này giúp chuyển pyruvate thành acetyl CoA. Các hợp chất này thay vào đó sẽ tạo ra các thể ketone [ketone bodies] và được gọi là các hợp chất tạo ketone [ketogenic]).
III. Các phản ứng
Bảy phản ứng đường phân là có thể đảo ngược và được sử dụng trong sự tổng hợp glucose từ lactate và pyruvate. Tuy nhiên, 3 phản ứng đường phân là không thể đảo ngược và phải được đi tránh qua bởi 4 phản ứng thay thế mà thuận lợi về mặt năng lượng cho sự tổng hợp glucose. Các phản ứng không thể đảo ngược này cùng với nhau là các phản ứng duy nhất đối với quá trình tân tạo đường, và được mô tả bên dưới.
A. Sự carboxyl hóa pyruvate
Rào chắn thứ nhất cần phải vượt qua trong sự tổng hợp glucose từ pyruvate là sự chuyển đổi không thể đảo ngược trong quá trình đường phân của PEP thành pyruvate bởi pyruvate kinase (PK). Trong quá trình tân tạo đường, pyruvate được carboxyl hóa bởi pyruvate carboxylase (PC) thành OAA, chất mà được chuyển thành PEP bởi PEP-carboxykinase (PEPCK) (Hình 3).
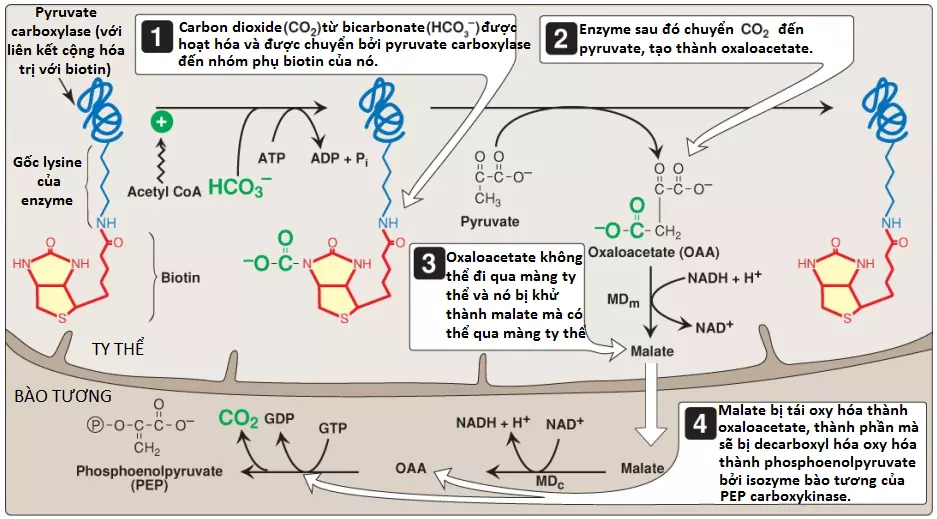
1. Biotin: PC cần coenzyme biotin liên kết cộng hóa trị với nhóm ε-amino của một gốc lysine trong enzyme (xem Hình 3). Sự thủy phân ATP điều khiển sự hình thành của một trung gian enzyme-biotin-carbon dioxide (CO2), trung gian này sau đó carboxyl hóa pyruvate để hình thành nên OAA. (Chú ý: HCO3– sẽ cung cấp CO2). Phản ứng PC xảy ra trong ty thể của các tế bào gan và thận và có hai mục đích: cho phép sự sản xuất PEP, một cơ chất quan trọng cho quá trình tân tạo đường, và cung cấp OAA mà có thể bổ sung lại các trung gian của chu trình TCA mà có thể trở nên cạn kiệt. Các tế bào cơ cũng chứa PC nhưng sử dụng sản phẩm OAA chỉ cho mục đích bổ sung và không tổng hợp glucose. (Chú ý: Protein mang pyruvate giúp di chuyển pyruvate từ bào tương vào trong ty thể).
PC là một trong số nhiều carboxylases mà cần biotin. Các carboxylases khác bao gồm acetyl CoA carboxylase, propionyl CoA carboxylase và methylcrotonyl CoA carboxylase.
2. Sự điều hòa dị lập thể: PC được hoạt hóa dị lập thể bởi acetyl CoA. Các mức acetyl CoA tăng lên trong ty thể truyền tín hiệu cho một trạng thái chuyển hóa mà trong đó sự tăng tổng hợp OAA là cần thiết. Ví dụ, điều này xảy ra trong suốt quá trình nhịn ăn (fasting), khi OAA được sử dụng cho quá trình tân tạo đường trong gan và các thận. Ngược lại, ở các mức acetyl CoA thấp, PC bị bất hoạt nhiều và pyruvate chủ yếu bị oxy hóa bởi PDHC thành acetyl CoA, thành phần mà có thể được oxy hóa tiếp bởi chu trình TCA.
B. Sự vận chuyển oxaloacetate đến bào tương
Khi quá trình tân tạo đường tiếp tục, OAA phải được chuyển thành PEP bởi PEPCK. Sự sản xuất PEP trong bào tương cần sự vận chuyển OAA ra khỏi ty thể. Tuy nhiên, không có chất vận chuyển OAA trong màng trong ty thể và OAA đầu tiên được khử thành malate bởi malate dehydrogenase ty thể (malate dehydrogenase – MD). Malate được vận chuyển vào trong bào tương và tái oxy hóa thành OAA bởi MD bào tương khi nicotinamide adenine dinucleotide (NAD+) được khử thành NADH (xem Hình 3). NADH được sử dụng trong sự khử 1,3-bisphosphoglycerate thành glyceraldehyde 3-phosphate bởi glyceraldehyde 3-phosphate dehydrogenase, một phản ứng chung đối với cả quá trình đường phân và quá trình tân tạo đường. (Chú ý: Khi có nhiều, lactate bị oxy hóa thành pyruvate khi NAD+ bị khử. Pyruvate được vận chuyển vào trong ty thể và được carboxyl hóa bởi PC thành OAA, chất mà có thể được chuyển thành PEP bởi isozyme PEPCK của ty thể. PEP được vận chuyển đến bào tương. OAA cũng có thể được chuyển thành aspartate mà được vận chuyển vào trong bào tương).
C. Sự decarboxyl hóa oxaloacetate của bào tương
OAA được decarboxyl hóa và phosphoryl hóa thành PEP trong bào tương bởi PEPCK. Phản ứng được điều khiển bởi sự thủy phân của guanosine triphosphate (GTP) (xem Hình 3). Các hoạt động kết hợp của PC và PEPCK cung cấp một con đường chuyển hóa thích hợp về mặt năng lượng từ pyruvate thành PEP. PEP sau đó được tác động bởi các phản ứng của quá trình đường phân diễn ra theo hướng ngược lại cho đến khi nó trở thành fructose 1,6-bisphosphate.
Sự bắt cặp của quá trình carboxyl hóa với quá trình decarboxyl hóa sẽ điều khiển các phản ứng mà nếu không có các quá trình này thì sẽ không thuận lợi về mặt năng lượng. Cách này cũng được sử dụng trong sự tổng hợp acid béo (fatty acid synthesis).
D. Sự dephosphoryl hóa fructose 1,6-bisphosphate
Sự thủy phân fructose 1,6-bisphosphate bởi fructose 1,6-bisphosphatase, được phát hiện trong gan và các thận, bỏ qua phản ứng phosphofructokinase-1 (PFK-1) không thể đảo ngược của quá trình đường phân và cung cấp một con đường thuận lợi về mặt năng lượng cho sự hình thành của fructose 6-phosphate (Hình 4). Phản ứng này là một vị trí điều hòa quan trọng của quá trình tân tạo đường.

1. Sự điều hòa bởi các mức năng lượng nội bào: Fructose 1,6-bisphosphate bị ức chế bởi một sự tăng lên trong tỷ số của adenosine monophosphate (AMP) với ATP, được gọi là tỷ số AMP với ATP, tỷ số này truyền tín hiệu về một trạng thái năng lượng thấp trong tế bào. Ngược lại, các mức AMP thấp và ATP cao kích thích quá trình tân tạo đường, một con đường yêu cầu năng lượng.
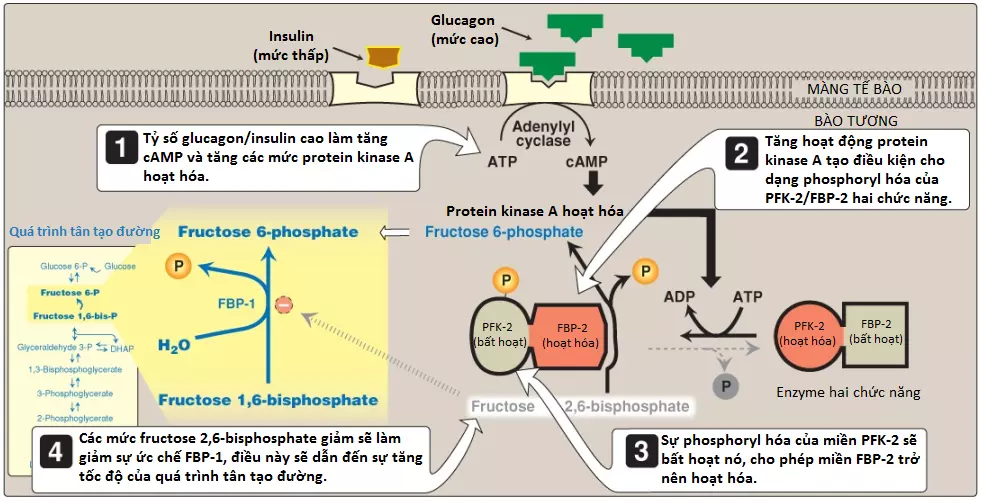
2. Sự điều hòa bởi fructose 2,6-bisphosphate: Fructose 1,6-bisphosphatase bị ức chế bởi fructose 2,6-bisphosphate, một chất tác động dị lập thể mà nồng độ của nó bị ảnh hưởng bởi tỷ số insulin/glucagon. Khi mức glucagon cao, chất tác động thì không được sản xuất bởi PFK-2 của gan và vì thế, phosphatase được hoạt hóa (Hình 5). (Chú ý: Các tín hiệu mà ức chế [năng lượng thấp, mức fructose 2,6-bisphosphate cao] hay hoạt hóa [năng lượng cao, mức fructose 2,6-bisphosphate thấp] quá trình tân tạo đường thì sẽ có tác động trái ngược lên quá trình đường phân, cung cấp sự kiểm soát qua lại của các con đường mà tổng hợp và oxy hóa glucose).
E. Sự dephosphoryl hóa glucose 6-phosphate
Sự thủy phân glucose 6-phosphate bởi glucose 6-phosphatase bỏ qua phản ứng hexokinase/glucokinase không thể đảo ngược và cung cấp một con đường thuận lợi về mặt năng lượng cho sự hình thành của glucose tự do (Hình 6). Gan là cơ quan chủ yếu sản xuất glucose tự do từ glucose 6-phosphate. Quá trình này yêu cầu một phức hợp của 2 protein chỉ được tìm thấy bên trong mô diễn ra sự tân tạo đường: glucose 6-phosphate translocase, thành phần mà vận chuyển glucose qua màng lưới nội chất và glucose 6-phosphatase , thành phần loại bỏ phosphate, tạo ra glucose tự do (xem Hình 6). Các protein màng lưới nội chất này cũng được cần cho bước cuối cùng của sự thoái hóa glycogen.

Các bệnh tích trữ glycogen Ia và Ib, được gây ra bởi các sự suy giảm tương ứng trong phosphatase và translocase được đặc trưng bởi sự hạ đường huyết khi nhịn ăn nghiêm trọng bởi vì glucose tự do thì không thể được tạo ra từ quá trình tân tạo đường hoặc quá trình thoái hóa glycogen. Các chất vận chuyển chuyên biệt chịu trách nhiệm cho sự di chuyển glucose tự do vào trong bào tương và sau đó vào trong máu.
F. Tổng hợp các phản ứng của quá trình đường phân và tân tạo đường
Trong số 11 phản ứng cần để chuyển pyruvate thành glucose tự do thì 7 phản ứng là được xúc tác bởi các enzyme đường phân có thể đảo ngược được (Hình 7). 3 phản ứng không thể đảo ngược, được xúc tác bởi hexokinase/glucokinase, PFK-1 và PK thì được né tránh bởi các phản ứng xúc tác bởi glucose 6-phosphatase, fructose 1,6-bisphosphate, PC và PEPCK. Trong quá trình tân tạo đường, các sự cân bằng của các phản ứng đường phân có thể đảo ngược thì sẽ được thúc đẩy về phía tổng hợp glucose như là kết quả của sự hình thành về bản chất là không thể đảo ngược của PEP, fructose 6-phosphate và glucose bởi các enzymes tân tạo đường. (Chú ý: Phân tích tỷ lượng của quá trình tân tạo đường từ hai phân tử pyruvate sẽ thấy được sự bắt cặp của sự phân tách 6 liên kết phosphate cao năng lượng và sự oxy hóa của 2 NADH với sự hình thành của một phân tử glucose [xem Hình 7]).

IV. Sự điều hòa
Sự điều hòa liên tục của quá trình tân tạo đường được xác định chủ yếu bởi mức tuần hoàn của glucagon và bởi sự có mặt của các cơ chất tân tạo đường. Ngoài ra, các thay đổi thích nghi chậm trong lượng enzyme là do một sự thay đổi trong tốc độ tổng hợp hay thoái hóa enzyme hay cả hai. (Chú ý: Sự kiểm soát hormone của hệ thống điều hòa chuyển hóa glucose sẽ được trình bày trong các bài viết tiếp theo của mình nhé).
A. Glucagon
Hormone peptide đến từ các tế bào tiểu đảo α của tụy này kích thích quá trình đường phân bởi 3 cơ chế:
1. Các sự thay đổi trong các chất tác động dị lập thể: Glucagon làm hạ mức fructose 2,6-bisphosphate gan, từ đó làm hoạt hóa fructose 1,6-bisphosphatase và ức chế PFK-1, bằng cách đó, tạo điều kiện thuận lợi cho quá trình tân tạo đường diễn ra mạnh hơn quá trình đường phân (xem Hình 5).
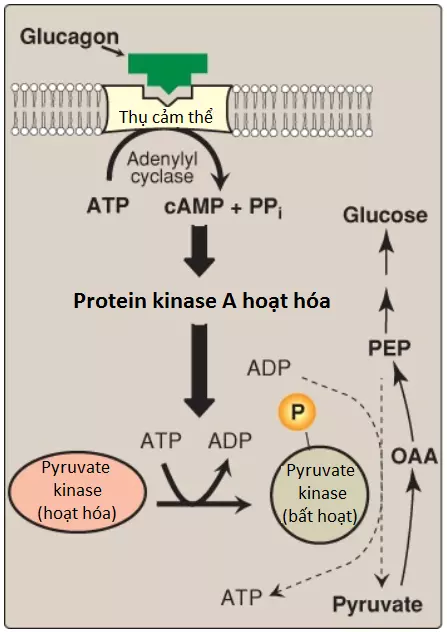
2. Sự chỉnh sửa cộng hóa trị hoạt động enzyme: Glucagon liên kết với thụ cảm thể bắt cặp protein G của nó và thông qua một sự tăng lên của mức AMP vòng (cyclic AMP – cAMP) và hoạt động của protein kinase A phụ thuộc cAMP, giúp kích thích sự chuyển đổi của PK gan thành dạng bất hoạt (dạng phosphoryl hóa của nó). Điều này làm giảm sự chuyển đổi PEP thành pyruvate, do đó, có tác động làm điều hướng PEP vào trong quá trình tân tạo đường (Hình 8).
3. Sự cảm ứng của quá trình tổng hợp enzyme: Glucagon làm tăng sự phiên mã của gene PEPCK thông qua yếu tố dịch mã protein liên kết thành phần đáp ứng cAMP (CREB), bằng cách đó, làm tăng cường sự có mặt của enzyme này khi các mức cơ chất của nó tăng trong suốt quá trình nhịn ăn. Cortisol, một glucocorticoid, cũng làm tăng sự biểu hiện của gene, ngược lại insulin làm giảm sự biểu hiện của gene.
B. Sự có mặt của cơ chất
Sự có mặt của các tiền thân tân tạo đường, đặc biệt là các amino acids tân tạo đường, sẽ ảnh hưởng đáng kể lên tốc độ của sự tổng hợp glucose. Các mức insulin giảm sẽ tạo điều kiện thuận lợi cho sự huy động các amino acids từ protein cơ để cung cấp các khung carbon cho quá trình tân tạo đường. Các coenzymes ATP và NADH cần cho quá trình tân tạo đường chủ yếu được cung cấp bởi sự oxy hóa acid béo.
C. Sự hoạt hóa dị lập thể bởi acetyl CoA
Sự hoạt hóa dị lập thể của PC gan bởi acetyl CoA xảy ra trong suốt quá trình nhịn ăn. Do sự tăng thủy phân TAG trong mô mỡ, gan sẽ chứa nhiều acid béo. Tốc độ hình thành acetyl CoA bởi quá trình oxy hóa β của các acid béo này sẽ vượt quá khả năng của gan trong việc oxy hóa nó thành CO2 và nước. Kết quả là acetyl CoA tích tụ lại và hoạt hóa PC. (Chú ý: Acetyl CoA ức chế PDHC [bằng cách hoạt hóa PDH kinase]. Vì thế, hợp chất đơn này có thể đưa pyruvate vào trong quá trình tân tạo đường và đi khỏi chu trình TCA [Hình 9]).

D. Sự ức chế dị lập thể bởi AMP
Fructose 1,6-bisphosphatase bị ức chế bởi AMP, một hợp chất mà sẽ hoạt hóa PFK-1. Điều này tạo ra sự điều hòa qua lại của quá trình đường phân và quá trình tân tạo đường mà được thấy trước đây với fructose 2,6-bisphosphate. Vì thế, tăng AMP sẽ kích thích các con đường tạo năng lượng và ức chế các con đường yêu cầu năng lượng.
V. Tổng hợp bài viết
- Các tiền thân tân tạo đường (gluconeogenic precursors) bao gồm glycerol giải phóng trong suốt quá trình thủy phân TAG trong mô mỡ, lactate được giải phóng bởi các tế bào thiếu ty thể và bởi cơ đang luyện tập thể thao, và các α-keto acids (như α-ketoglutarate và pyruvate) có nguồn gốc từ sự chuyển hóa amino acid tân tạo đường (Hình 10).
- Bảy phản ứng trong quá trình đường phân là có thể đảo ngược và được sử dụng cho quá trình tân tạo đường trong gan và các thận.
- Ba phản ứng, được xúc tác bởi PK, PFK-1 và glucokinase/hexokinase là không thể đảo ngược về mặt sinh lý và phải được đi tránh qua.
- Pyruvate được chuyển thành OAA và sau đó thành PEP bởi PC và PEPCK.
- PC cần biotin và ATP và được hoạt hóa về mặt dị lập thể bởi acetyl CoA và PEPCK cần GTP.
- Fructose 1,6-bisphosphate được chuyển thành fructose 6-phosphate bởi fructose 1,6-bisphosphatase. Enzyme này bị ức chế bởi một tỷ số AMP/ATP cao và bởi fructose 2,6-bisphosphate, chất hoạt hóa dị lập thể chủ yếu của quá trình đường phân.
- Glucose 6-phosphate bị dephosphoryl hóa thành glucose bởi glucose 6-phosphatase. Enzyme này của màng lưới nội chất xúc tác bước cuối cùng trong quá trình tân tạo đường và trong sự thoái hóa glycogen. Sự suy giảm của enzyme này gây ra tình trạng hạ đường huyết khi nhịn ăn (fasting hypoglycemia) nghiêm trọng.
Các bạn có thể xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn có thể xem bài viết trước tại đây: https://docsachxyz.com/chu-trinh-tricarboxylic-acid-va-phuc-hop-pyruvate-dehydrogenase/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!







{kind=link}