





























I. Tổng quan
Vi sinh vật gây bệnh được định nghĩa là vi sinh vật có khả năng gây bệnh (độc lực). Một số vi sinh vật là vi sinh vật gây bệnh rõ ràng, trong khi những vi sinh vật khác (phần lớn) thường không gây bệnh. Một vi sinh vật có thể xâm nhập vào một cá thể mà không gây ra các triệu chứng rõ ràng, có thể phát hiện được. Sự xuất hiện của các bệnh nhiễm trùng không triệu chứng như vậy có thể được nhận biết thông qua sự hiện diện của vi sinh vật hoặc sự hiện diện của kháng thể chống lại các vi sinh vật đó ở bệnh nhân. Một số bệnh nhiễm trùng dẫn đến trạng thái “tiềm ẩn”, nghĩa là vi sinh vật đang ở trạng thái không hoạt động nhưng có thể được kích hoạt lại khi các triệu chứng phục hồi. Hơn nữa, một số tác nhân gây bệnh chỉ gây bệnh trong một số điều kiện nhất định (ví dụ, được đưa vào một vị trí cơ thể bình thường vô trùng hoặc nhiễm trùng ở vật chủ bị suy giảm miễn dịch). Hình 1 tóm tắt một số thuật ngữ được sử dụng để mô tả sự đa dạng của các bệnh nhiễm trùng.

II. Các cơ chế bệnh sinh vi khuẩn
Mặc dù các cơ chế cụ thể của quá trình nhiễm trùng khác nhau giữa các tác nhân gây bệnh khác nhau, nhưng phương pháp mà vi khuẩn gây bệnh có thể được chia thành một số giai đoạn chung (Hình 2). Khả năng gây bệnh của một vi sinh vật phụ thuộc vào mức độ thành công của nó trong việc hoàn thành một số hoặc tất cả các giai đoạn này. Các thuật ngữ “độc lực” và “khả năng gây bệnh” thường được sử dụng thay thế cho nhau. Tuy nhiên, độc lực có thể được định lượng bằng số lượng vi sinh vật cần thiết để gây bệnh ở 50% số sinh vật tiếp xúc với tác nhân gây bệnh (ID50, trong đó I = Nhiễm trùng [Infectious] và D = Liều lượng [Dose]) hoặc để giết chết 50% động vật thí nghiệm (LD50, trong đó L = Gây tử vong [Lethal]). Số lượng sinh vật cần thiết để gây bệnh rất khác nhau giữa các vi khuẩn gây bệnh. Ví dụ, <100 Shigella gây tiêu chảy bằng cách lây nhiễm đường tiêu hóa (GI), trong khi liều gây bệnh của Salmonella là ~100,000 sinh vật nhưng thay đổi tùy thuộc vào thành phần của sản phẩm thực phẩm bị nhiễm. Liều gây bệnh của một loại vi khuẩn cũng phụ thuộc vào các yếu tố độc lực của nó. Xác suất xảy ra một bệnh truyền nhiễm phụ thuộc vào cả liều lượng và độc lực của vi sinh vật gây bệnh, bên cạnh sức mạnh của phản ứng miễn dịch chống lại nhiễm trùng của vật chủ.

A. Các yếu tố độc lực
Các yếu tố độc lực là những đặc điểm của vi khuẩn làm tăng khả năng gây bệnh của nó, tức là các đặc tính cho phép vi sinh vật tự thiết lập và nhân lên trên hoặc bên trong một vật chủ cụ thể. Một số bước quan trọng hơn trong quá trình lây nhiễm được xem xét dưới đây.
1. Xâm nhập vào vật chủ: Bước đầu tiên của quá trình lây nhiễm là vi sinh vật xâm nhập vào vật chủ qua một trong một số cổng: qua đường hô hấp, đường tiêu hóa hoặc đường tiết niệu-sinh dục hoặc qua da bị cắt, thủng hoặc bỏng. Sau khi xâm nhập thành công, tác nhân gây bệnh phải vượt qua nhiều cơ chế phòng vệ khác nhau của vật chủ trước khi có thể thiết lập được. Những cơ chế này bao gồm thực bào, môi trường axit của dạ dày và đường tiết niệu-sinh dục, và nhiều loại enzyme thủy phân và phân giải protein có trong nước bọt, dạ dày và ruột non. Vi khuẩn có lớp vỏ polysaccharide bên ngoài (ví dụ: Streptococcus pneumoniae và Neisseria meningitidis) có nhiều khả năng sống sót hơn sau những cơ chế phòng vệ chính của vật chủ này.
2. Bám dính vào tế bào vật chủ: Một số vi khuẩn (ví dụ, Escherichia coli; xem phần sau) sử dụng pili (còn gọi là fimbriae) để bám dính vào bề mặt tế bào vật chủ. Neisseriae có cấu trúc tương tự (xem phần sau). Các vi khuẩn khác có các phân tử bám dính bề mặt tế bào hoặc thành tế bào đặc biệt kỵ nước mà cho phép chúng bám dính vào màng tế bào vật chủ. Trong mỗi trường hợp, sự bám dính làm tăng độc lực bằng cách ngăn vi khuẩn bị chất nhầy mang đi hoặc bị rửa trôi khỏi các cơ quan có dòng chảy chất lỏng đáng kể, chẳng hạn như đường tiết niệu và đường tiêu hóa. Sự bám dính cũng cho phép mỗi tế bào vi khuẩn bám dính tạo thành một vi khuẩn lạc. Một ví dụ nổi bật về tầm quan trọng của sự bám dính là Neisseria gonorrhoeae, trong đó các chủng không có pili không gây bệnh (xem phần sau).
3. Khả năng xâm lấn: Vi khuẩn xâm lấn là vi khuẩn có thể xâm nhập vào tế bào vật chủ hoặc xuyên qua bề mặt niêm mạc, lây lan từ vị trí nhiễm trùng ban đầu. Sự xâm lấn được tạo điều kiện thuận lợi bởi các enzyme của vi khuẩn, bao gồm collagenase và hyaluronidase. Các enzyme này phân hủy các thành phần của chất nền ngoại bào, giúp vi khuẩn dễ dàng tiếp cận bề mặt tế bào vật chủ hơn. Nhiều tác nhân gây bệnh vi khuẩn biểu hiện các protein màng được gọi là “invasins” tương tác với các thụ thể tế bào vật chủ, do đó gây ra các chuỗi tín hiệu dẫn đến sự hấp thụ vi khuẩn bằng thực bào cảm ứng. Xâm lấn thường được theo sau bởi tình trạng viêm, có thể là viêm mủ (bao gồm sự hình thành mủ) hoặc viêm hạt (có tổn thương viêm dạng nốt), tùy thuộc vào sinh vật. Mủ của tình trạng viêm mủ chủ yếu chứa bạch cầu trung tính, trong khi tổn thương hạt chứa nguyên bào sợi, tế bào lympho và đại thực bào.
4. Cô lập sắt: Sắt là chất dinh dưỡng thiết yếu đối với hầu hết các vi khuẩn. Để có được sắt cần thiết cho sự phát triển, vi khuẩn sản xuất các hợp chất liên kết sắt, được gọi là siderophores. Các hợp chất này bắt giữ sắt từ vật chủ bằng cách tạo phức, sau đó siderophore được ferrate hóa liên kết với các thụ thể đặc hiệu trên bề mặt vi khuẩn. Sắt được vận chuyển tích cực vào vi khuẩn, tại đó nó được kết hợp vào các hợp chất thiết yếu như cytochrome. Các loài Neisseria gây bệnh là ngoại lệ ở chỗ chúng không sản xuất siderophore mà thay vào đó sử dụng các protein liên kết sắt của vật chủ, chẳng hạn như transferrin và lactoferrin, làm nguồn sắt. Chúng làm như vậy bằng cách biểu hiện các thụ thể chuyên dụng liên kết với các protein của vật chủ này và loại bỏ sắt cho sự nội hóa.
5. Các yếu tố độc lực ức chế thực bào: Cấu trúc chống thực bào quan trọng nhất là lớp vỏ bên ngoài thành tế bào, chẳng hạn như ở S. pneumoniae và N. meningitidis. Nhóm yếu tố chống thực bào thứ hai là các protein thành tế bào của cầu khuẩn gram dương, chẳng hạn như protein A của tụ cầu khuẩn và protein M của liên cầu khuẩn nhóm A (xem các phần sau).
6. Độc tố vi khuẩn: Một số vi khuẩn gây bệnh bằng cách sản xuất các chất độc hại, trong đó có hai loại chung: ngoại độc tố và nội độc tố. Ngoại độc tố là các protein do cả vi khuẩn gram dương và gram âm tiết ra. Ngược lại, nội độc tố, đồng nghĩa với lipopolysaccharide (LPS), không được tiết ra mà là một thành phần không thể thiếu của thành tế bào vi khuẩn gram âm.
a. Ngoại độc tố: Các độc tố tiết ra bao gồm một số chất độc nhất được biết đến. Người ta ước tính rằng chỉ cần 1 µg ngoại độc tố uốn ván cũng có thể giết chết một người trưởng thành. Protein ngoại độc tố thường có hai thành phần polypeptide (Hình 3). Một thành phần chịu trách nhiệm liên kết protein với tế bào vật chủ và một thành phần chịu trách nhiệm về tác dụng độc hại. Trong một số trường hợp, mục tiêu chính xác của độc tố đã được xác định. Ví dụ, độc tố bạch hầu là một loại enzyme ngăn chặn quá trình tổng hợp protein. Nó thực hiện điều này bằng cách gắn một nhóm adenosine diphosphate-ribosyl vào yếu tố kéo dài protein EF-2 của người, do đó vô hiệu hóa yếu tố này (xem phần sau). Hầu hết các ngoại độc tố đều bị bất hoạt nhanh chóng khi đun nóng ở nhiệt độ vừa phải (60°C), các ngoại lệ đáng chú ý là độc tố ruột của tụ cầu và độc tố chịu nhiệt (heat-stable toxin – ST) của E. coli. Ngoài ra, xử lý bằng formaldehyde loãng sẽ phá hủy hoạt tính độc hại của hầu hết các ngoại độc tố nhưng không ảnh hưởng đến tính kháng nguyên của chúng. Các độc tố bị bất hoạt bằng formaldehyde, được gọi là các toxoids, rất hữu ích trong việc chế tạo vắc-xin (xem phần sau). Trong nhiều trường hợp, protein ngoại độc tố được mã hóa bởi các gen mang trên plasmid hoặc thực khuẩn thể ôn hòa. Một ví dụ là ngoại độc tố bạch hầu được mã hóa bởi gen độc tố của thực khuẩn thể ôn hòa có thể gây tan Corynebacterium diphtheriae. Các chủng C. diphtheriae mang loại thực khuẩn này là chủng gây bệnh, trong khi các chủng không có loại thực khuẩn này là chủng không gây bệnh.
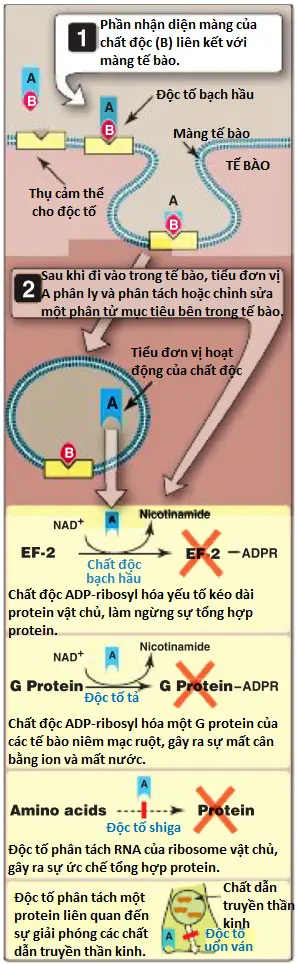
b. Nội độc tố: Đồng nghĩa với LPS, nội độc tố là thành phần chịu nhiệt của màng ngoài của vi khuẩn gram âm (nhưng không có trong gram dương). Chúng được giải phóng vào hệ tuần hoàn của vật chủ trong quá trình phát triển của vi khuẩn hoặc sau khi tế bào vi khuẩn bị ly giải. LPS bao gồm polysaccharide, bao gồm các tiểu đơn vị đường lặp lại (kháng nguyên O), nhô ra khỏi bề mặt tế bào bên ngoài, một polysaccharide lõi và một thành phần lipid gọi là lipid A được tích hợp vào lá ngoài của màng ngoài. Phần lipid A chịu trách nhiệm về độc tính của phân tử này. Các tác động sinh lý chính của nội độc tố hoặc LPS là sốt, sốc, hạ huyết áp và huyết khối, được gọi chung là sốc nhiễm trùng. Những tác động này được tạo ra gián tiếp thông qua hoạt hóa đại thực bào, với việc giải phóng cytokine, hoạt hóa bổ thể và kích hoạt chuỗi đông máu. Tử vong có thể là kết quả của suy đa cơ quan. Việc loại bỏ vi khuẩn gây bệnh bằng kháng sinh ban đầu có thể làm trầm trọng thêm các triệu chứng bằng cách gây ra sự giải phóng nội độc tố đột ngột và ồ ạt vào hệ tuần hoàn. Mặc dù vi khuẩn gram dương không chứa LPS, peptidoglycan thành tế bào và axit teichoic của chúng có thể gây ra hội chứng sốc tương tự như hội chứng do LPS gây ra, nhưng thường không nghiêm trọng bằng.
B. Bệnh sinh qua trung gian vật chủ
Bệnh sinh của nhiều bệnh nhiễm trùng do vi khuẩn là do phản ứng của vật chủ chứ không phải do các yếu tố của vi khuẩn. Các ví dụ điển hình về bệnh sinh qua phản ứng của vật chủ được thấy trong các bệnh như nhiễm trùng huyết do vi khuẩn gram âm, bệnh lao và bệnh phong củ. Tổn thương mô trong các bệnh nhiễm trùng này là do nhiều cytokine khác nhau được giải phóng từ tế bào lympho, đại thực bào và bạch cầu đa nhân tại vị trí nhiễm trùng hoặc trong máu. Phản ứng của vật chủ thường rất mạnh đến mức phá hủy các mô của vật chủ, cho phép vi khuẩn còn lại tăng sinh.
C. Biến đổi pha và kháng nguyên
Một tác nhân gây bệnh thành công phải trốn tránh được các yếu tố hệ thống miễn dịch của vật chủ mà nhận ra các kháng nguyên bề mặt của vi khuẩn. Một “chiến lược” trốn tránh quan trọng được một số tác nhân gây bệnh sử dụng là thay đổi các kháng nguyên bề mặt của nó. Điều này được thực hiện bằng một số cơ chế. Một cơ chế, được gọi là biến dị pha (phase variation), là khả năng có thể đảo ngược về mặt di truyền của một số vi khuẩn để tắt và bật biểu hiện của các gen mã hóa cho các kháng nguyên bề mặt. Một cơ chế thứ hai, được gọi là biến dị kháng nguyên, liên quan đến việc sửa đổi gen cho một kháng nguyên bề mặt được biểu hiện bằng cách tái tổ hợp di truyền với một trong nhiều trình tự DNA không biểu hiện khác nhau. Theo cách này, kháng nguyên bề mặt được biểu hiện có thể có nhiều cấu trúc kháng nguyên khác nhau (xem phần sau).
Các bạn có thể xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn có thể xem bài viết trước tại đây: https://docsachxyz.com/he-vi-sinh-vat-binh-thuong/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!







{kind=link}