





























I. Tổng quan về chuyển hóa
Trong các bài viết trước, các phản ứng enzyme riêng lẻ đã được phân tích để giải thích các cơ chế của xúc tác. Tuy nhiên trong các tế bào, các phản ứng này hiếm khi xảy ra riêng lẻ. Thay vào đó, chúng được tổ chức thành các trình tự phản ứng gồm nhiều bước được gọi là các con đường chuyển hóa, như đường phân chẳng hạn (Hình 1). Trong một con đường chuyển hóa, sản phẩm của một phản ứng đóng vai trò như là một cơ chất của phản ứng tiếp theo. Hầu hết các con đường chuyển hóa có thể được phân loại thành dị hóa (catabolic) (thoái hóa (degradative)) hay đồng hóa (anabolic) (tổng hợp (synthetic)). Các con đường dị hóa thoái hóa các phân tử phức tạp, như proteins, polysaccharides và lipids, thành một số phân tử đơn giản (như carbon dioxide, ammonia và nước). Các con đường đồng hóa hình thành nên các sản phẩm phức tạp cuối cùng từ các tiền thân đơn giản, ví dụ, sự tổng hợp polysaccharide glycogen từ glucose. Các con đường chuyển hóa khác nhau có thể giao nhau, hình thành nên một mạng lưới các phản ứng hóa học tổng hợp và có mục đích. Sự chuyển hóa là tổng hợp tất cả các thay đổi hóa học xảy ra trong một tế bào, một mô hay toàn bộ cơ thể. Các chất chuyển hóa là các sản phẩm trung gian của chuyển hóa. Các bài viết tiếp theo của mình sẽ tập trung vào các con đường chuyển hóa trung tâm mà liên quan đến sự tổng hợp và thoái hóa carbohydrates, lipids và amino acids.

A. Sơ đồ chuyển hóa
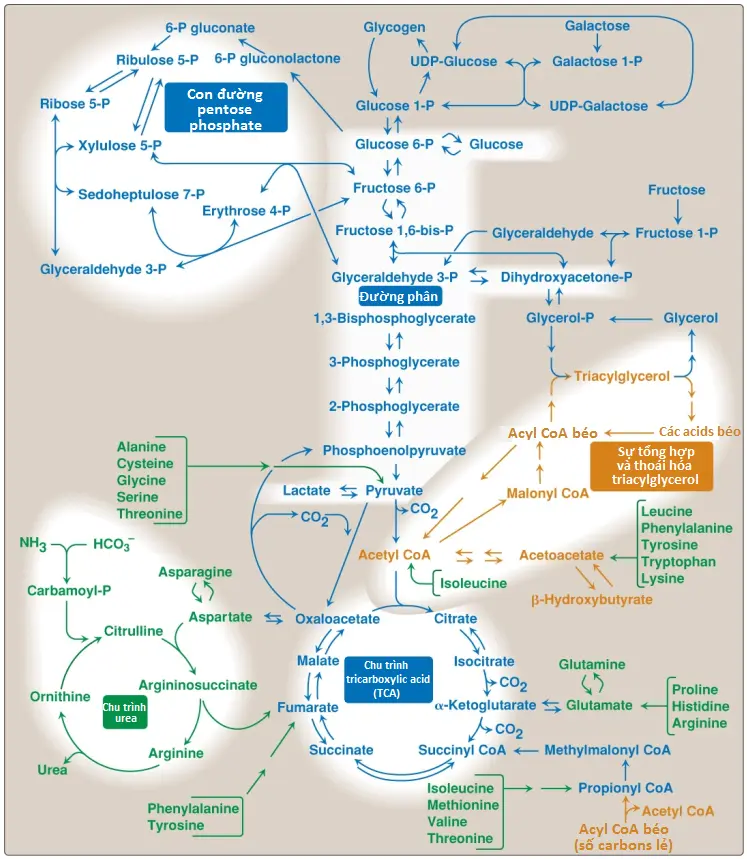
Sự chuyển hóa được hiểu tốt nhất bằng cách xem xét các con đường chuyển hóa thành phần của nó. Mỗi con đường đều bao gồm các trình tự đa enzyme và mỗi enzyme cuối cùng sẽ thực hiện các vai trò xúc tác hay điều hòa quan trọng. Một sơ đồ chuyển hóa chứa các con đường chuyển hóa trung tâm quan trọng của sự chuyển hóa năng lượng được thể hiện trên Hình 2. “Bức tranh tổng quan” này của sự chuyển hóa thì hữu ích trong việc tìm được các sự kết nối giữa các con đường chuyển hóa, hình dung sự di chuyển có mục đích của các chất chuyển hóa và mô tả tác động lên dòng các chất trung gian nếu như một con đường chuyển hóa bị ức chế hay ngăn chặn, ví dụ, một sự suy giảm enzyme do thuốc hay do di truyền. Trong suốt các bài viết tiếp theo của mình thì mỗi con đường chuyển hóa được bàn đến sẽ mô tả lại như là một phần của sơ đồ chuyển hóa lớn được thể hiện trên Hình 2.
B. Các con đường dị hóa
Các phản ứng dị hóa đóng vai trò trong việc tích trữ năng lượng ở dạng ATP từ sự thoái hóa của các phân tử nguyên liệu giàu năng lượng. Sự hình thành ATP bằng sự thoái hóa các phân tử phức tạp xảy ra trong 3 giai đoạn như được thể hiện trong Hình 3. (Chú ý: Các con đường dị hóa thường là oxy hóa và yêu cầu các coenzymes oxy hóa như nicotinamide adenine dinucleotide [NAD+]). Sự dị hóa cũng cho phép các phân tử trong chế độ ăn hoặc các phân tử chất dinh dưỡng tích trữ trong các tế bào được chuyển thành các chất cơ bản cần cho sự tổng hợp các phân tử phức tạp. Sự dị hóa do đó sẽ được mô tả như là một quá trình chuyển đổi mà trong đó nhiều loại phân tử được chuyển dạng thành một số sản phẩm chung cuối cùng.

1. Sự thủy phân của các phân tử phức tạp: Trong giai đoạn đầu tiên, các phân tử phức tạp được phân tách thành các chất cơ bản. Ví dụ, các proteins được thoái hóa thành các amino acids, các polysaccharides được thoái hóa thành các monosaccharides và các chất béo (triacylglycerols) được thoái hóa thành các acids béo và glycerol.
2. Sự chuyển đổi của các thành phần cơ bản thành các trung gian đơn giản: Trong giai đoạn thứ hai, các cấu trúc cơ bản phong phú này sẽ được thoái hóa hơn nữa thành acetyl coenzyme A (CoA) và một số phân tử đơn giản khác. Một ít năng lượng được tích trữ dưới dạng ATP nhưng với lượng nhỏ so với năng lượng được sản xuất ra trong suốt giai đoạn thứ ba của quá trình dị hóa.
3. Sự oxy hóa acetyl CoA: Chu trình tricarboxylic acid (TCA) (xem các bài viết sau của mình) là con đường chung cuối cùng trong sự oxy hóa các phân tử nguyên liệu mà tạo ra acetyl CoA. Sự oxy hóa acetyl CoA tạo ra các lượng lớn ATP thông qua sự phosphoryl hóa oxy hóa khi các electrons đi từ NADH và flavin adenine dinucleotide (FADH2) đến oxygen ([O2], xem lại bài viết trước của mình nhé).
C. Các con đường đồng hóa
Ngược lại với sự dị hóa, sự đồng hóa là một quá trình khác mà trong đó một số tiền thân sinh tổng hợp (như các amino acids) hình thành nên nhiều loại sản phẩm phức tạp chuỗi dài (như các proteins [Hình 4]). Các phản ứng đồng hóa yêu cầu năng lượng (thu năng lượng), nhìn chung là được cung cấp bởi sự thủy phân của ATP thành adenosine diphosphate (ADP) và phosphate vô cơ (Pi). (Chú ý: Các phản ứng dị hóa tạo ra năng lượng [tỏa năng lượng]). Các phản ứng đồng hóa thường liên quan đến các sự khử hóa học mà trong đó năng lượng khử thường được cung cấp bởi chất cho electron NADPH (NADH được phosphoryl hóa, xem các bài viết tiếp theo của mình để hiểu).

II. Sự điều hòa chuyển hóa
Các con đường chuyển hóa phải được phối hợp để sự sản xuất năng lượng hay sự tổng hợp các sản phẩm cuối cùng đáp ứng các nhu cầu của tế bào. Hơn thế nữa, các tế bào riêng lẻ đóng vai trò như là một phần của một tập hợp các mô tương tác, chứ không phải là tách biệt với nhau. Vì thế, một hệ thống liên hệ phức tạp đã được tiến hóa để phối hợp các chức năng của cơ thể. Các tín hiệu điều hòa mà truyền cho một tế bào riêng rẽ về trạng thái chuyển hóa của cơ thể nói chung bao gồm các hormones, các chất dẫn truyền thần kinh và sự có sẵn của các chất dinh dưỡng. Các tín hiệu điều hòa này cuối cùng sẽ ảnh hưởng đến các tín hiệu được tạo ra bên trong tế bào (Hình 5).

A. Sự giao tiếp nội bào
Tốc độ của một con đường chuyển hóa có thể đáp ứng các tín hiệu điều hòa mà xuất phát từ bên trong tế bào. Ví dụ, tốc độ chuyển hóa có thể bị ảnh hưởng bởi sự sẵn có của các cơ chất, sự ức chế sản phẩm, hay các sự thay đổi trong mức các chất hoạt hóa dị lập thể hay các chất ức chế dị lập thể. Các tín hiệu nội bào này thường tạo ra các sự đáp ứng nhanh và quan trọng cho sự điều hòa trong khoảng thời gian ngắn của sự chuyển hóa.
B. Sự giao tiếp gian bào
Khả năng đáp ứng với các tín hiệu gian bào là cần thiết cho sự phát triển và tồn tại của các sinh vật. Sự truyền tín hiệu giữa các tế bào cung cấp một sự phối hợp rộng rãi của chuyển hóa và thường tạo ra một đáp ứng như một sự thay đổi trong biểu hiện gene mà xuất hiện chậm hơn so với các tín hiệu nội bào. Sự giao tiếp giữa các tế bào có thể được điều hòa, ví dụ, bởi sự hình thành của các liên kết khe, cho phép sự giao tiếp trực tiếp giữa các bào tương của các tế bào lân cận. Tuy nhiên, đối với sự chuyển hóa năng lượng, con đường giao tiếp quan trọng nhất là sự truyền tín hiệu hóa học giữa các tế bào bằng các hormones qua đường máu hay bằng các chất dẫn truyền thần kinh.
C. Các thụ cảm thể liên kết protein và các hệ thống truyền tin thứ hai
Các hormones và các chất dẫn truyền thần kinh có thể được cho là các tín hiệu và các thụ cảm thể (receptor) của chúng đóng vai trò như là các thành phần phát hiện tín hiệu. Các thụ cảm thể là các proteins thường được vùi trong màng tế bào của các tế bào đích. Chúng đáp ứng với một ligand (chất hóa học) liên kết với chúng bằng cách khởi động một chuỗi các phản ứng mà cuối cùng sẽ tạo ra các đáp ứng nội bào đặc hiệu. Nhiều thụ cảm thể mà giúp điều hòa sự chuyển hóa thì được liên kết với các proteins liên kết GTP nội bào (intracellular GTP-binding proteins) được gọi là các proteins G và được biết đến như là các thụ cảm thể bắt cặp protein G (GPCRs). Loại thụ cảm thể này điều hòa sự sản xuất của các phân tử được gọi là các chất truyền tin thứ hai, chúng được đặt tên như vậy là do chúng can thiệp giữa chất truyền tin ngoại bào ban đầu (các chất dẫn truyền thần kinh hay hormone) và tác động nội bào cuối cùng. Các chất truyền tin thứ hai là một phần của chuỗi các sự kiện kế tiếp mà giúp chuyển đổi liên kết ligand thành một đáp ứng nội bào.
Hai trong số các số hệ thống nhận diện chất truyền tin thứ hai thường gặp nhất được điều hòa bởi các proteins G là hệ thống phospholipase C mà liên quan đến canxi và hệ thống phosphatidylinositol và hệ thống adenylyl cyclase (adenylate cyclase), đặc biệt quan trọng trong điều hòa các con đường của sự chuyển hóa trung gian. Cả hai hệ thống đều được khởi đầu bằng sự liên kết của các ligand hormone như epinephrine hay glucagon với một GPCR vùi trong màng bào tương của tế bào đích mà sẽ đáp ứng với hormone.
GPCRs được đặc trưng bởi một miền liên kết với ligand ở ngoại bào, 7 xoắn α xuyên màng và một miền nội bào mà tương tác với các trimer không đồng nhất G proteins gồm 3 tiểu đơn vị α, β và γ (xem Hình 6). (Chú ý: Insulin, một chất điều hòa quan trọng khác của chuyển hóa thì không truyền tín hiệu qua GPCRs nhưng thay vào đó nó sẽ tác động qua một thụ cảm thể liên quan với hoạt động của tyrosine kinase [xem các bài viết tiếp theo của mình nhé]).

D. Adenylyl cyclase
Sự liên kết của ligand hormone với một số GPCRs, bao gồm các thụ cảm thể β-adrenergic và α2-adrenergic, sẽ khơi mào cho một sự tăng lên hoặc giảm đi trong hoạt động của adenylyl cyclase. Đây là một enzyme liên kết với màng giúp chuyển ATP thành 3′,5′-adenosine monophosphate (AMP vòng, hay cAMP) khi hoạt hóa.
1. Các proteins điều hòa phụ thuộc guanosine triphosphate hay proteins G: Tác động của GPCR liên kết với ligand và được hoạt hóa lên sự hình thành của chất truyền tin thứ hai thì được điều hòa bởi các trimers proteins G không đồng nhất (gồm các tiểu đơn vị α, β và γ) được tìm thấy trên mặt trong của màng bào tương. Các proteins G được đặt tên như thế là bởi vì tiểu đơn vị α của nó liên kết với guanosine triphosphate (GTP) khi được hoạt hóa. Ở dạng bất hoạt của một G protein, tiểu đơn vị α được liên kết với GDP (Hình 7). Sự liên kết ligand gây ra một sự thay đổi cấu hình trong thụ cảm thể, khơi mào cho sự thay thế của GDP này bằng GTP. Dạng liên kết GTP của tiểu đơn vị α sẽ phân ly khỏi các tiểu đơn vị βγ và di chuyển đến enzyme adenylyl cyclase liên kết với màng, tác động lên hoạt động enzyme của nó. Nhiều phân tử protein Gα hoạt hóa được hình thành bởi một thụ cảm thể được kích hoạt. (Chú ý: Khả năng của một hormone hoặc một chất dẫn truyền thần kinh kích thích hoặc ức chế adenylyl cyclase phụ thuộc vào loại Gα protein mà liên kết với receptor. Một loại, được định danh là Gs, sẽ kích thích adenylyl cyclase [xem Hình 7], ngược lại, Gi sẽ ức chế adenylyl cyclase [không được thể hiện]).
Adenylyl cyclase hoạt hóa chuyển adenosine triphosphate (ATP) thành chất truyền tin thứ hai cAMP hay adenosine monophosphate vòng. cAMP sau đó hoạt hóa serine/threonine protein kinase được gọi là protein kinase A (PKA), được mô tả bên dưới. Các hoạt động của phức hợp Gα-GTP thì có thời gian tồn tại ngắn bởi vì Gα có một hoạt tính GTPase cố hữu, gây ra sự thủy phân nhanh của GTP thành GDP. Điều này gây ra sự bất hoạt của Gα, sự phân ly của nó khỏi adenylyl cyclase và sự tái kết hợp của nó với dimer βγ.
Các độc tố từ Vibrio cholerae (vi khuẩn tả) và Bordetella pertussis (vi khuẩn ho gà) gây ra sự hoạt hóa không thích hợp của adenylyl cyclase thông qua sự chỉnh sửa cộng hóa trị (ADP-ribosyl hóa (ADP-ribosylation)) của các proteins G khác nhau mà tương tác với adenylyl cyclase. Với độc tố vi khuẩn tả, hoạt động GTPase của Gαs bị ức chế trong các tế bào ruột. Với ho gà, độc tố vi khuẩn ho gà bất hoạt Gαi trong các tế bào đường hô hấp. Kết quả trong cả hai trường hợp là sự tăng hoạt động của adenylyl cyclase và sự sản xuất quá mức chất truyền tin thứ hai, cAMP.
2. Protein kinases: Bước tiếp theo trong hệ thống chất truyền tin thứ hai cAMP là sự hoạt hóa của một họ enzymes được gọi là protein kinases phụ thuộc cAMP, bao gồm PKA, như được thể hiện trong Hình 8. cAMP hoạt hóa PKA bằng cách liên kết với 2 tiểu đơn vị điều hòa của nó, gây ra sự giải phóng của hai tiểu đơn vị hoạt động có hoạt tính xúc tác của nó. PKA hoạt động là một serine/threonine kinase bởi vì nó thực hiện chức năng chuyển phosphate từ ATP đến các gốc serine hay threonine đặc hiệu của các cơ chất protein chuyên biệt của nó. Các proteins được phosphoryl hóa có thể tác động trực tiếp lên các kênh ion của tế bào hay nếu các enzymes thì có thể trở nên hoạt hóa hay ức chế. (Chú ý: Không phải tất cả các loại protein kinases là phụ thuộc cAMP, ví dụ như protein kinase C, được hoạt hóa trong đáp ứng với sự truyền tín hiệu của phospholipase C, là phụ thuộc canxi).

3. Protein phosphatases: Các nhóm phosphate thêm vào trong các proteins bởi các protein kinases thì được loại bỏ bởi các phosphoprotein phosphatases, các enzymes mà phân tách các esters phosphate bằng sự thủy phân (xem Hình 8). Các hoạt động của phosphatases đảm bảo rằng các sự thay đổi trong hoạt động protein được thúc đẩy bởi sự phosphoryl hóa là không phải vĩnh viễn.
4. Sự thủy phân cAMP: cAMP được thủy phân nhanh chóng thành 5’-AMP bởi cAMP phosphodiesterase mà giúp phân tách liên kết 3′,5′-phosphodiester vòng. 5’-AMP thì không phải là một phân tử truyền tín hiệu nội bào. Vì thế, các tác động làm tăng cAMP qua trung gian chất dẫn truyền thần kinh và qua trung gian hormone thì được kết thúc một cách nhanh chóng nếu như tín hiệu ngoại bào bị loại bỏ. (Chú ý: cAMP phosphodiesterase bị ức chế bởi caffeine, một dẫn xuất methylxanthine).
Các bạn có thể xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn có thể xem bài viết trước tại đây: https://docsachxyz.com/gioi-thieu-ve-carbohydrates/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!







{kind=link}