





























I. Tổng quan con đường pentose phosphate
Con đường pentose phosphate (pentose phosphate pathway), còn được gọi là chuyển hướng hexose monophosphate (hexose monophosphate shunt), giúp cung cấp ribose 5-phosphate cho sự sinh tổng hợp của các nucleotides và nó có vai trò quan trọng vì là nguồn nicotinamide adenine dinucleotide phosphate (NADPH) chính của cơ thể, đây là một chất khử sinh hóa. NADPH là nguồn các đương lượng khử của tế bào cho sự sinh tổng hợp của các acid béo và cholesterol và cho sự khử hydrogen peroxide (H2O2) mà được hình thành trong quá trình đáp ứng với stress oxy hóa và là một sản phẩm phụ của quá trình chuyển hóa hiếu khí. Glucose 6-phosphate dehydrogenase (G6PD) xúc tác cho bước giới hạn tốc độ đầu tiên của con đường chuyển hóa; sự di truyền liên kết nhiễm sắc thể X của tình trạng thiếu hụt G6PD gây ra sự thiếu hụt NADPH, đặc biệt là trong các tế bào hồng cầu, làm cho chúng dễ bị phân giải trong đáp ứng với stress oxy hóa. Con đường này không sản xuất hay tiêu thụ ATP.
Các phản ứng của con đường này xảy ra trong bào tương và bao gồm một giai đoạn oxy hóa không thể đảo ngược, theo sau bởi một loạt các sự chuyển đổi qua lại của đường-phopshate mà có thể đảo ngược (Hình 1). Trong giai đoạn oxy hóa, carbon thứ nhất của phân tử glucose 6-phosphate được giải phóng dưới dạng carbon dioxide (CO2) và một đường pentose-phosphate cùng với hai NADPHs khử được tạo ra. Tốc độ và hướng của các phản ứng có thể đảo ngược được xác định bởi sự cung cấp của và nhu cầu đối với các trung gian của con đường chuyển hóa. Con đường pentose phosphate cũng tạo ra ribose 5-phosphate, cần cho sự sinh tổng hợp nucleotide (xem thêm các bài viết sau này của mình) và cung cấp một cơ chế cho sự chuyển đổi của các đường pentose thành các trung gian triose và hexose của quá trình đường phân.

II. Các phản ứng oxy hóa không thể đảo ngược
Phần oxy hóa của con đường pentose phosphate bao gồm 3 phản ứng không thể đảo ngược mà dẫn đến sự hình thành của ribose 5-phosphate, CO2 và hai phân tử NADPH cho mỗi phân tử glucose 6-phosphate oxy hóa (Hình 2). Phần này của con đường chuyển hóa thì đặc biệt quan trọng trong gan, các tuyến vú đang tạo sữa và mô mỡ đối với sự sinh tổng hợp phụ thuộc NADPH của các acid béo (xem thêm các bài viết sau của mình); trong các tinh hoàn, các buồng trứng, nhau thai và vỏ thượng thận đối với sự sinh tổng hợp các hormones steroid phụ thuộc NADPH (xem thêm các bài viết sau của mình); và trong các tế bào hồng cầu đối với sự khử glutathione phụ thuộc NADPH.

A. Glucose 6-phosphate dehydrogenase
Glucose 6-phosphate dehydrogenase (G6PD) xúc tác sự oxy hóa glucose 6-phosphate thành 6-phosphogluconolactone khi coenzyme NADP+ bị khử thành NADPH. Phản ứng khởi động này là bước bước cam kết, giới hạn tốc độ và điều hòa (committed, rate-limiting, and regulated step) của con đường chuyển hóa. NADPH là một chất ức chế tranh chấp mạnh mẽ của G6PD và tỷ số của NADPH/NADP+ là đủ cao để ức chế đáng kể enzyme dưới hầu hết các điều kiện chuyển hóa. Tuy nhiên, với nhu cầu tăng lên đối với NADPH thì tỷ số NADPH/NADP+ giảm và dòng vật chất qua con đường chuyển hóa tăng lên trong đáp ứng với hoạt động tăng cường của G6PD. Nên chú ý rằng là insulin tăng cường điều hòa biểu hiện gene của G6PD và dòng vật chất qua con đường tăng lên trong trạng thái hấp thu (xem thêm các bài viết sau của mình).
B. Sự hình thành của ribulose 5-phosphate
6-phosphogluconolactone được thủy phân bởi 6-phosphogluconolactone hydrolase trong bước thứ hai. Sự decarboxyl hóa oxy hóa của sản phẩm, là 6-phosphogluconate, được xúc tác bởi 6-phosphogluconate dehydrogenase. Bước không thể đảo ngược thứ ba này tạo ra ribulose 5-phosphate, là một đường pentose-phosphate, CO2 (từ carbon thứ nhất của glucose) và một phân tử NADPH thứ hai (xem Hình 2).
III. Các phản ứng không phải oxy hóa có thể đảo ngược
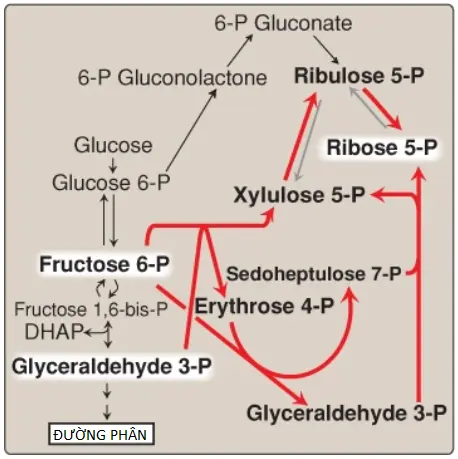
Các phản ứng không phải oxy hóa của con đường pentose phopshate xảy ra trong tất cả các loại tế bào tổng hợp nucleotides và nucleic acid. Các phản ứng này xúc tác sự chuyển đổi qua lại của các đường chứa 3 đến 7 carbons (xem Hình 2). Các phản ứng có thể đảo ngược này cho phép ribulose 5-phosphate được sản xuất bởi phần oxy hóa của con đường chuyển hóa được chuyển thành ribose 5-phosphate cần cho sự tổng hợp nucleotide (xem thêm các bài viết sau của mình) hoặc thành các trung gian của quá trình đường phân, là fructose 6-phosphate và glyceraldehyde 3-phosphate.
Nhiều tế bào thực hiện các phản ứng sinh tổng hợp khử có một nhu cầu đối với NADPH lớn hơn so với ribose 5-phosphate. Trong trường hợp này, transketolase giúp chuyển các đơn vị 2 carbon trong một phản ứng cần thiamine pyrophosphate (TPP) và transaldolase giúp chuyển các đơn vị 3 carbon, chuyển đổi ribulose 5-phosphate được tạo ra như là một sản phẩm cuối cùng của giai đoạn oxy hóa thành glyceraldehyde 3-phosphate và fructose 6-phosphate. Ngược lại, khi nhu cầu đối với ribose cho các nucleotides và các nucleic acids lớn hơn nhu cầu của NADPH thì các phản ứng không phải oxy hóa có thể cung cấp ribose 5-phosphate từ glyceraldehyde 3-phosphate và fructose 6-phosphate trong sự vắng mặt của các bước oxy hóa (Hình 3).
Ngoài transketolase, TPP thì được cần bởi các phức hợp đa enzyme pyruvate dehydrogenase (các bạn xem lại bài viết trước của mình nhé), , α-ketoglutarate dehydrogenase của chu trình tricarboxylic acid (xem lại bài viết trước của mình) và α-keto acid phân nhánh dehydrogenase (branched-chain α-keto acid dehydrogenase) của quá trình dị hóa amino acid phân nhánh (xem thêm các bài viết sau của mình).
IV. Các tác dụng của NADPH
Coenzyme NADPH khác so với nicotinamide adenine dinucleotide (NADH) chỉ bởi sự có mặt của nhóm phosphate trên một trong các đơn vị ribose (Hình 4). Sự thay đổi có vẻ là nhỏ này trong cấu trúc cho phép NADPH tương tác với các enzymes đặc hiệu NADPH mà có các vai trò nhất định trong tế bào. Ví dụ, trong bào tương của các tế bào gan, tỷ số NADP+/NADPH ở trạng thái ổn định là khoảng 0.1, điều này tạo điều kiện thuận lợi cho việc sử dụng NADPH trong các phản ứng sinh tổng hợp khử. Điều này tương phản với tỷ số NAD+/NADH cao (khoảng 1,000), điều này tạo điều kiện thuận lợi cho vai trò oxy hóa đối với NAD+. Phần này tóm lược một số chức năng đặc hiệu quan trọng của NADPH trong các phản ứng sinh tổng hợp khử và các phản ứng giải độc.

A. Sự sinh tổng hợp khử
Giống như NADH, NADPH có thể được xem là một phân tử cao năng lượng. Tuy nhiên, các electrons của NADPH được sử dụng cho sự sinh tổng hợp khử hơn là cho chuỗi truyền electron như được thể hiện với NADH (xem lại bài viết trước của mình). Trong các sự chuyển dạng chuyển hóa của con đường pentose phosphate, một phần năng lượng của glucose 6-phosphate được tích trữ trong NADPH, một phân tử có điện thế khử âm (xem lại bài viết trước của mình), vì thế, nó có thể được sử dụng trong các phản ứng cần một chất cho electron như sự tổng hợp acid béo (xem thêm bài viết sau của mình), sự tổng hợp cholesterol và sự tổng hợp hormone steroid (xem thêm các bài viết sau của mình).
B. Sự khử H2O2
H2O2 là một thành viên của một họ các gốc oxygen phản ứng (reactive oxygen species – ROS) mà được hình thành từ sự khử một phần oxygen phân tử, O2 (Hình 5A). Các hợp chất này được tạo thành một cách liên tục như là các sản phẩm phụ của chuyển hóa hiếu khí, qua các phản ứng với các thuốc và các chất độc môi trường hoặc khi mức các chất chống oxy hóa bị giảm xuống, tất cả tạo ra tình trạng của stress oxy hóa (oxidative stress). Các trung gian oxygen phản ứng cao này có thể gây ra sự tổn thương hóa học nghiêm trọng đối với DNA, các proteins và các lipids không bão hòa và có thể dẫn đến sự chết tế bào. ROS có liên quan trong một số các quá trình bệnh lý, bao gồm tổn thương tái tưới máu (reperfusion injury), ung thư, bệnh viêm nhiễm và lão hóa (aging). Tế bào có một vài cơ chế bảo vệ mà làm giảm thiểu khả năng gây hại của các hợp chất này. ROS cũng có thể được tạo ra trong quá trình tiêu diệt các vi sinh vật bởi các tế bào bạch cầu (xem phần tiếp theo của bài viết).

1. Các enzymes mà xúc tác các phản ứng chống oxy hóa: Glutathione khử (G-SH) là một tripeptide-thiol (γ-glutamylcysteinylglycine) có trong hầu hết các tế bào, có thể khử độc H2O2 về mặt hóa học (Hình 5B). Phản ứng này, được xúc tác bởi glutathione peroxidase, hình thành nên glutathione oxy hóa (G-S-S-G), thành phần không có các thuộc tính bảo vệ nữa. Tế bào tái tạo G-SH trong một phản ứng được xúc tác bởi glutathione reductase, sử dụng NADPH như là một nguồn các đương lượng khử. Vì thế, NADPH cung cấp các electrons một cách gián tiếp cho sự khử H2O2 (Hình 6). Các enzymes khác như superoxide dismutase và catalase, xúc tác sự chuyển đổi của các ROS khác nhau thành các sản phẩm không gây hại (xem Hình 5B). Như là một nhóm, các enzymes này đóng vai trò như là một hệ thống phòng thủ giúp bảo vệ cơ thể khỏi các tác động gây hại của ROS.

2. Các chất hóa học chống oxy hóa: Một số chất khử nội bào như ascorbate hay vitamin C, vitamin E và β-carotene là có khả năng khử và bằng cách đó, giải độc ROS trong phòng thí nghiệm. Sự tiêu thụ các thức ăn giàu các hợp chất chống oxy hóa này có liên quan với sự giảm nguy cơ của các loại ung thư nhất định cũng như là giảm tần suất của các vấn đề sức khỏe mạn tính khác. Vì thế, thật muốn đưa ra ý kiến rằng là các tác động của các hợp chất này thì phần nào là biểu hiện khả năng của chúng trong ngăn chặn tác động gây hại của ROS. Tuy nhiên, các thử nghiệm lâm sàng với các chất chống oxy hóa dưới dạng bổ sung trong chế độ ăn thì không cho thấy các tác động có lợi rõ ràng. Trong trường hợp bổ sung trong chế độ ăn với β-carotene thì tỷ lệ ung thư phổi ở những người hút thuốc tăng lên thay vì là giảm đi. Vì thế, các tác động tăng cường sức khỏe của các trái cây và rau củ trong chế độ ăn dường như phản ánh một sự tương tác phức tạp giữa nhiều hợp chất xuất hiện một cách tự nhiên mà không giống với việc tiêu thụ các hợp chất chống oxy hóa riêng lẻ (xem thêm các bài viết tiếp theo của mình).
C. Hệ thống cytochrome P450 monooxygenase

Monooxygenases (oxidases chức năng hỗn hợp) tích hợp một nguyên tử từ O2 vào trong một cơ chất (tạo một nhóm hydroxyl), với nguyên tử còn lại được khử thành nước (H2O). Trong hệ thống cytochrome P450 (CYP) monooxygenase, NADPH cung cấp các đương lượng khử cần bởi chuỗi các phản ứng này (Hình 7). Hệ thống này thực hiện các chức năng khác nhau trong 2 vị trí riêng biệt trong các tế bào. Phản ứng tổng quan xúc tác bởi một enzyme CYP là:

trong đó R có thể là một steroid, thuốc hoặc các chất hóa học khác. Các enzymes CYP thực sự là một siêu họ của các monooxygenases chứa heme liên quan mà tham gia vào trong nhiều phản ứng. P450 trong tên phản ánh khả năng hấp thu bước sóng tối đa ở 450 nm bởi protein.
1. Hệ thống ty thể (mitochondrial system): Một chức năng quan trọng của hệ thống CYP monooxygenase được tìm thấy liên quan với màng trong ty thể là sự sinh tổng hợp của các hormone steroid. Trong các mô tạo steroid như nhau thai, buồng trứng, tinh hoàn và vỏ thượng thận, nó được sử dụng để hydroxyl hóa các trung gian trong sự chuyển đổi của cholesterol thành các hormone steroid, một quá trình mà làm cho các hợp chất kỵ nước này dễ tan trong nước hơn (xem các bài viết sau của mình). Gan sử dụng cùng hệ thống này trong sự tổng hợp acid mật (xem các bài viết sau của mình) và sự hydroxyl hóa cholecalciferol thành 25-hydroxycholecalciferol (vitamin D3; xem các bài viết sau của mình) và thận sử dụng nó để hydroxyl hóa vitamin D3 thành dạng 1,25-dihydroxyl hóa hoạt động về mặt sinh học.
2. Hệ thống tiểu thể (microsomal system): Hệ thống CYP monooxygenase tiểu thể được tìm thấy liên quan với màng của lưới nội chất trơn, đặc biệt là trong gan, thực hiện chức năng chủ yếu trong sự giải độc các hợp chất ngoại lai hay “xenobiotics”. Các chất này bao gồm nhiều thuốc và các chất ô nhiễm khác nhau như các sản phẩm từ dầu mỏ và thuốc trừ sâu. Các enzymes CYP của hệ thống tiểu thể, ví dụ, CYP3A4, có thể được sử dụng để hydroxyl hóa các chất độc này (giai đoạn I). Mục đích của sự chỉnh sửa này là có hai mục đích. Đầu tiên, chính nó có thể hoạt hóa hoặc bất hoạt một thuốc và thứ hai, hình thành một chất độc dễ tan hơn, bằng cách đó, tăng cường sự bài tiết của nó trong nước tiểu hay phân. Tuy nhiên, thông thường, nhóm hydroxyl mới sẽ đóng vai trò như là một vị trí cho sự liên kết với một phân tử phân cực như glucuronic acid (xem các bài viết sau của mình), thành phần mà sẽ làm tăng đáng kể tính tan của hợp chất (giai đoạn II). Nên chú ý rằng là các sự đa hình (xem các bài viết tiếp theo của mình) trong các genes của các enzyme CYP có thể dẫn đến các sự khác nhau trong chuyển hóa thuốc.
Các bạn có thể xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn có thể xem bài viết trước tại đây: https://docsachxyz.com/chuyen-hoa-monosaccharide-va-disaccharide/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!







{kind=link}
Dạ cho em hỏi là những phản ứng không có chiều nghịch của con đường đường phân thì tại các phản ứng đó trong tân tạo đường sẽ giải phóng oxaloacetate đúng không ạ
Có các phản ứng chuyển pyruvate thành PEP qua trung gian oxaloacetate trong quá trình tân tạo đường là có tạo ra oxaloacetate (không phải tất cả các bước vì có 3 phản ứng ko thể đảo ngược của quá trình đường phân và được thay thế bởi 4 phản ứng “tránh né” trong quá trình tân tạo đường). Bạn đọc phần “A. Sự carboxyl hóa pyruvate” trở xuống là hiểu nhé !!!