





























III. Sự tổng hợp phospholipid
C. Phosphatidylserine
Sự tổng hợp PS ở các mô của động vật có vú được thực hiện bởi phản ứng trao đổi base, trong đó, ethanolamine của PE được trao đổi với serine tự do (xem Hình 6, phần 1). Phản ứng này, mặc dù có thể đảo ngược, nhưng được sử dụng chủ yếu để sản xuất PS cần cho sự tổng hợp màng. PS có tích điện âm toàn phần (xem các bài viết tiếp theo về vai trò của PS trong đông máu).
D. Phosphatidylinositol
Phosphatidylinositol (PI) được tổng hợp từ inositol tự do và CDP-DAG, như được thể hiện trong Hình 5 của phần 1. PI là một phospholipid không thông thường mà trong đó nó thường chứa stearic acid trên carbon 1 và arachidonic acid trên carbon 2 của glycerol. Vì thế, PI đóng vai trò như là một nơi chứa arachidonic acid trong các màng và vì thế, cung cấp cơ chất cho sự tổng hợp prostaglandin (PG) khi cần. Giống như PS, PI có tích điện âm toàn phần. (Chú ý: Có một sự không đối xứng trong thành phần phospholipid của màng tế bào. Ví dụ, PS và PI được tìm thấy chủ yếu trên lá trong của màng tế bào. Sự bất đối xứng này đạt được bởi các enzymes phụ thuộc ATP được gọi là “flippases” và “floppases”).
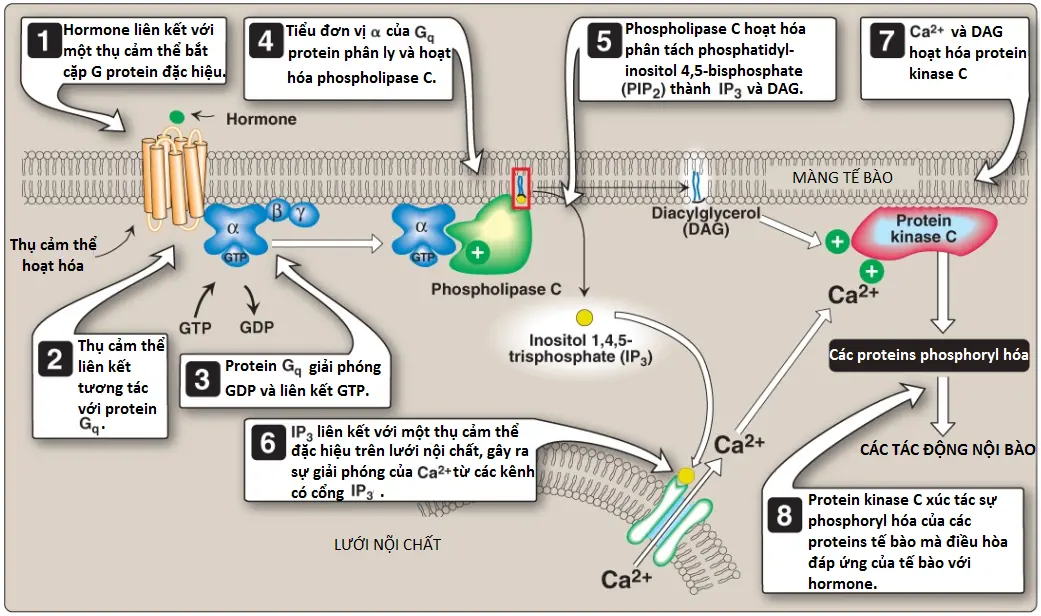
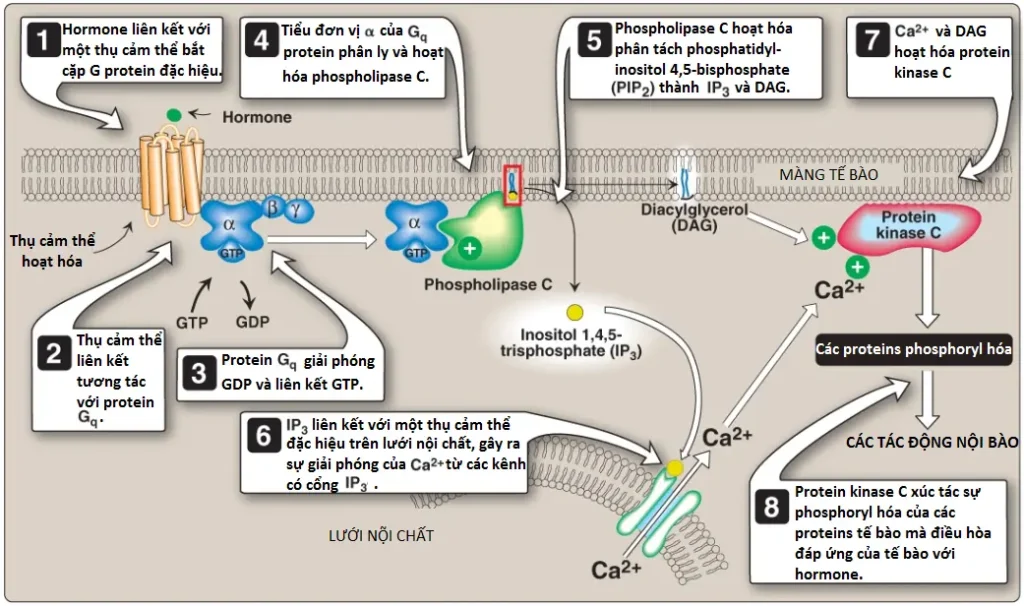
1. Vai trò trong sự truyền tín hiệu qua màng: Sự phopshoryl hóa của PI liên kết màng tạo ra polyphosphoinositides như phosphatidylinositol 4,5-bisphosphate ([PIP2]; Hình 7). Sự phân tách của PIP2 bởi phospholipase C xảy ra trong đáp ứng với sự liên kết của các chất dẫn truyền thần kinh, các hormones và các yếu tố tăng trưởng khác nhau với các thụ cảm thể bắt cặp với G protein (G protein–coupled receptors – GPCRs), như thụ cảm thể α1-adrenergic, trên màng tế bào và sự hoạt hóa của tiểu đơn vị α của Gq (Hình 8). Các sản phẩm của sự phân tách này, inositol 1,4,5-triphosphate (IP3) và DAG, điều hòa sự huy động của calcium nội bào và sự hoạt hóa của protein kinase C, mà đóng vai trò hợp lực để gây ra các đáp ứng tế bào chuyên biệt. Sự truyền tín hiệu qua màng vì thế được thực hiện.
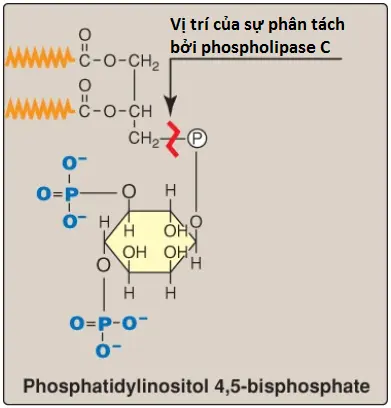
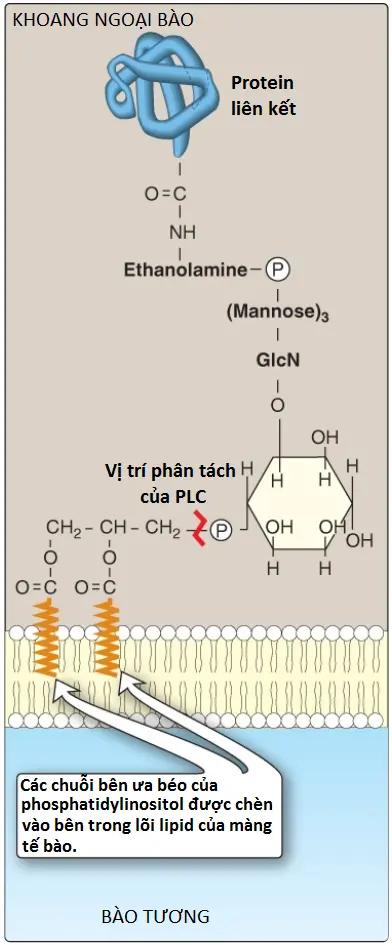
2. Vai trò trong việc neo giữ protein màng: Các protein chuyên biệt có thể được liên kết cộng hóa trị qua một cầu carbohydrate với PI liên kết màng (Hình 9). Ví dụ, lipoprotein lipase, một enzyme mà thoái hóa TAG trong các phần tử lipoproteins, được nối với các tế bào nội mô mao mạch bởi một kết nối glycosyl phosphatidylinositol (GPI). (Chú ý: Các proteins có liên kết GPI cũng được tìm thấy trong nhiều động vật nguyên sinh ký sinh, như trypanosomes và leishmania). Được nối với một lipid của màng (chứ không phải là một phần bên trong của màng) cho phép các protein liên kết GPI tăng tính di động sang bên trên mặt ngoại bào của màng bào tương. Protein có thể được phân tách khỏi chỗ nối của nó bởi hoạt động của phospholipase C (xem Hình 9). (Chú ý: Một thiếu hụt trong sự tổng hợp GPI trong các tế bào tạo máu gây ra bệnh huyết tán là đái huyết sắc tố kịch phát về đêm (paroxysmal nocturnal hemoglobinuria). Một vài trong số các proteins kết nối GPI giúp bảo vệ các tế bào máu khỏi hệ thống miễn dịch mà có vai trò nhận diện các chất ngoại lai như các virus và vi khuẩn trong cơ thể. Sự thiếu protein liên kết GPI trên các tế bào làm cho hồng cầu không còn được xem là “của cơ thể” nữa và dễ chịu sự phá hủy bởi sự phân giải tế bào qua trung gian bổ thể).
E. Phosphatidylglycerol và cardiolipin
Phosphatidylglycerol được tìm thấy với các nồng độ tương đối lớn trong các màng ty thể và là một tiền thân của cardiolipin (diphosphatidylglycerol). Nó được tổng hợp từ CDP-DAG và glycerol 3-phosphate. Cardiolipin (xem Hình 2, phần 1) được tổng hợp bởi sự chuyển của DAG 3-phosphate từ CDP-DAG đến một phân tử phosphatidylglycerol đã tồn tại từ trước.
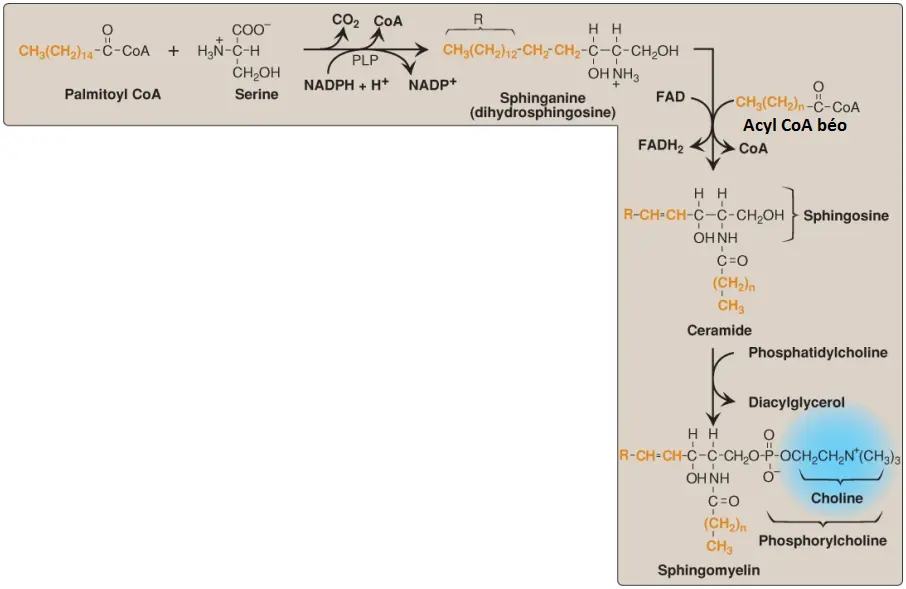
F. Sphingomyelin
Sphingomyelin, một phospholipid hình thành từ sphingosine, được tìm thấy trong các màng tế bào và trong bao myelin. Sự tổng hợp của sphingomyelin được thể hiện trong Hình 10. Nói ngắn gọn, palmitoyl CoA ngưng tụ với serine, khi CoA và nhóm carboxyl (dưới dạng CO2) bị mất đi. (Chú ý: Phản ứng này, giống với các phản ứng decarboxyl hóa liên quan với sự tổng hợp của PE từ PS và của các yếu tố điều hòa từ amino acids [như các catecholamines từ tyrosine; xem các bài viết sau], cần pyridoxal phosphate [một dẫn xuất của vitamin B6] như là một coenzyme). Sản phẩm bị khử trong một phản ứng cần nicotinamide adenine dinucleotide phosphate (NADPH) thành sphinganine (dihydrosphingosine). Sphinganine được acyl hóa ở nhóm amino với một trong số nhiều LCFA và sau đó giảm bão hòa để tạo ra ceramide, tiền thần gần nhất của sphingomyelin (và các sphingolipids khác, như được mô tả trong các phần sau).
Ceramides đóng một vai trò quan trọng trong việc duy trì hàng rào chống mất nước của da. Giảm các mức ceramide thì có liên quan với nhiều bệnh về da.
Phosphorylcholine từ PC được chuyển đến ceramide, tạo thành sphingomyelin và DAG. (Chú ý: Sphingomyelin của bao myelin chứa chủ yếu acid béo chuỗi dài như lignoceric acid và nervonic acid, ngược lại, chất xám của não có sphingomyelin mà chứa chủ yếu stearic acid).
IV. Sự thoái hóa của phospholipid
Sự thoái hóa của phosphoglycerides được thực hiện bởi phospholipases được tìm thấy trong tất cả các mô và dịch tụy. (Chú ý: Sự bàn luận về việc tiêu hóa phospholipid thì xem các bài viết trước nhé). Một số chất độc và nọc độc có hoạt tính phospholipase và một số vi khuẩn gây bệnh sản xuất phospholipases mà làm phân giải màng tế bào và cho phép sự lan rộng của nhiễm khuẩn. Sphingomyelin bị thoái hóa bởi phospholipase, sphingomyelinase của lysosome (xem phần B bên dưới).
A. Phosphoglycerides
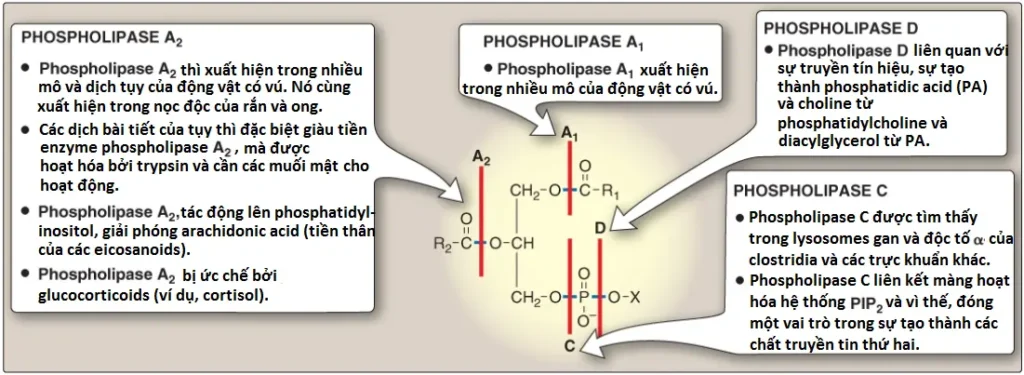
Phospholipases thủy phân các liên kết phosphodiester của phosphoglycerides, với mỗi enzyme phân tách phopspholipid ở một vị trí đặc hiệu. Các phospholipases chính được thể hiện trong Hình 11. (Chú ý: sự loại bỏ FA khỏi carbon 1 hoặc 2 của phosphoglyceride tạo ra một lysophosphoglyceride, là cơ chất cho lysophospholipases). Phospholipases giải phóng các phân tử mà có thể đóng vai trò như là các chất truyền tin thứ hai (như DAG và IP3) hoặc là cơ chất cho sự tổng hợp của các chất truyền tin (như arachidonic acid). Phospholipases không chỉ chịu trách nhiệm cho thoái hóa phospholipid mà còn tái cấu trúc chúng. Ví dụ, phospholipases A1 và A2 loại bỏ FA đặc hiệu khỏi các phospholipid liên kết màng, mà có thể được thay thế bởi FA khác nhờ sử dụng acyl CoA béo transferase. Cơ chế này được sử dụng như là một cách để tạo ra surfactant phổi độc nhất DPCC và để đảm bảo rằng carbon 2 của PI (và đôi khi là PC) được liên kết với arachidonic acid. (Chú ý: Hội chứng Barth, một rối loạn hiếm liên kết với NST X được đặc trưng bởi bệnh lý cơ tim, yếu cơ và giảm bạch cầu đa nhân trung tính, là kết quả của các khiếm khuyết trong sự tái cấu trúc cardiolipin).
B. Sphingomyelin
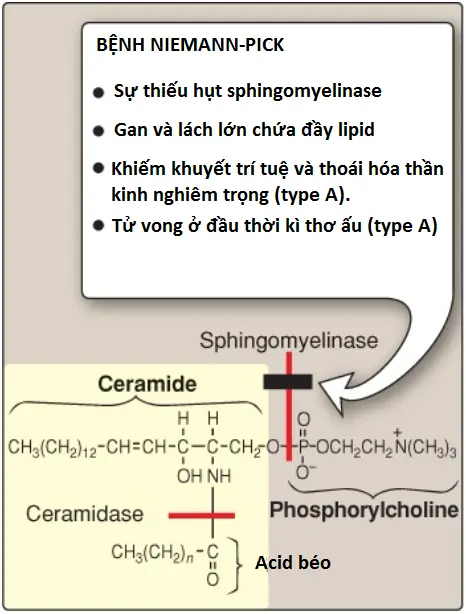
Sphingomyelin bị thoái hóa bởi sphingomyelinase, một enzyme của lysosome mà giúp loại bỏ phosphorylcholine, còn lại ceramide. Ceramide cuối cùng được phân tách bởi ceramidase thành sphingosine và một FA tự do (Hình 12). (Chú ý: Ceramide và sphingosine giải phóng điều hòa các con đường truyền tín hiệu, một phần bởi sự ảnh hưởng đến hoạt động của protein kinase C và vì thế, tác động đến sự phosphoryl hóa của các cơ chất protein của nó. Chúng cũng tăng cường sự chết theo chương trình tế bào). Bệnh Niemann-Pick (type A và B) là một rối loạn di truyền lặn trên nhiễm sắc thể thường được gây ra bởi sự mất khả năng thoái hóa sphingomyelin do một sự suy giảm của sphingomyelinase, một loại phospholipase C.
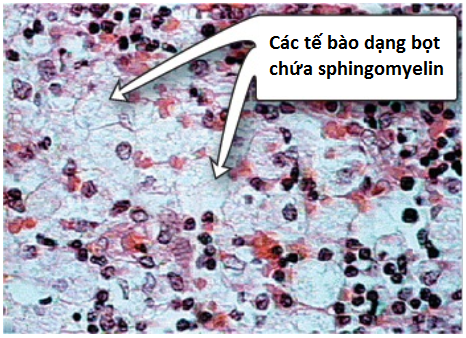
Trong thể sơ sinh nghiêm trọng (type A, cho thấy dưới 1% hoạt động enzyme bình thường), gan và lách là các vị trí lắng đọng lipids chủ yếu và vì thế, gan lách to phát triển. Lipid chứa chủ yếu là sphingomyelin mà không thể bị thoái hóa (Hình 13). Các đại thực bào của hệ thống võng nội mô trở nên ứ căng bởi sphingomyelin, điều này làm cho chúng có hình ảnh mô học giống như bọt. Những đứa trẻ sinh ra mắc bệnh tích trữ lysosome này sẽ trải qua sự thoái hóa thần kinh nhanh và tiến triển do sự tích tụ sphingomyelin trong hệ thống thần kinh trung ương. Kết quả, một điểm đỏ cherry trong điểm vàng của mắt phát triển do sự lắng đọng lipid và phù trong các tế bào hạch võng mạc. Những đứa trẻ này tử vong sớm trong đầu thời kỳ thơ ấu. Một biến thể ít nghiêm trọng hơn (type B, biến thể này cho thấy đến 10% hoạt động enzyme bình thường) với tuổi khởi phát trễ hơn và một thời gian sống sót dài hơn bởi vì có ít hoặc không có tổn thương mô thần kinh, nhưng các phổi, lách, gan và tủy xương thì bị ảnh hưởng, gây ra một dạng mạn tính của bệnh. Mặc dù bệnh Niemann-Pick xuất hiện ở tất cả các nhóm chủng tộc nhưng type A xảy ra với tần suất lớn hơn ở dân số Do Thái Ashkenazi. (Chú ý: Bệnh Niemann-Pick type C [NPC] gây ra bởi các đột biến ở NPC1 hay NPC2, là các genes quan trọng trong xử lý cholesterol được nhập bào và dẫn đến cả sự tích tụ cholesterol và sphingomyelin).
Các bạn xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn xem lại bài viết trước về phospholipid tại đây: https://docsachxyz.com/chuyen-hoa-phospholipid-glycosphingolipid-va-eicosanoid-phan-1/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!






{kind=link}