





























I. Tổng quan về chu trình
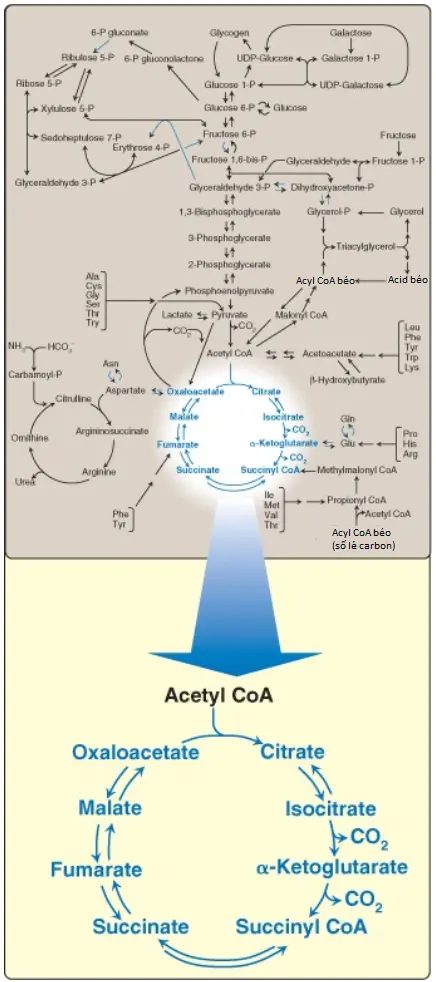
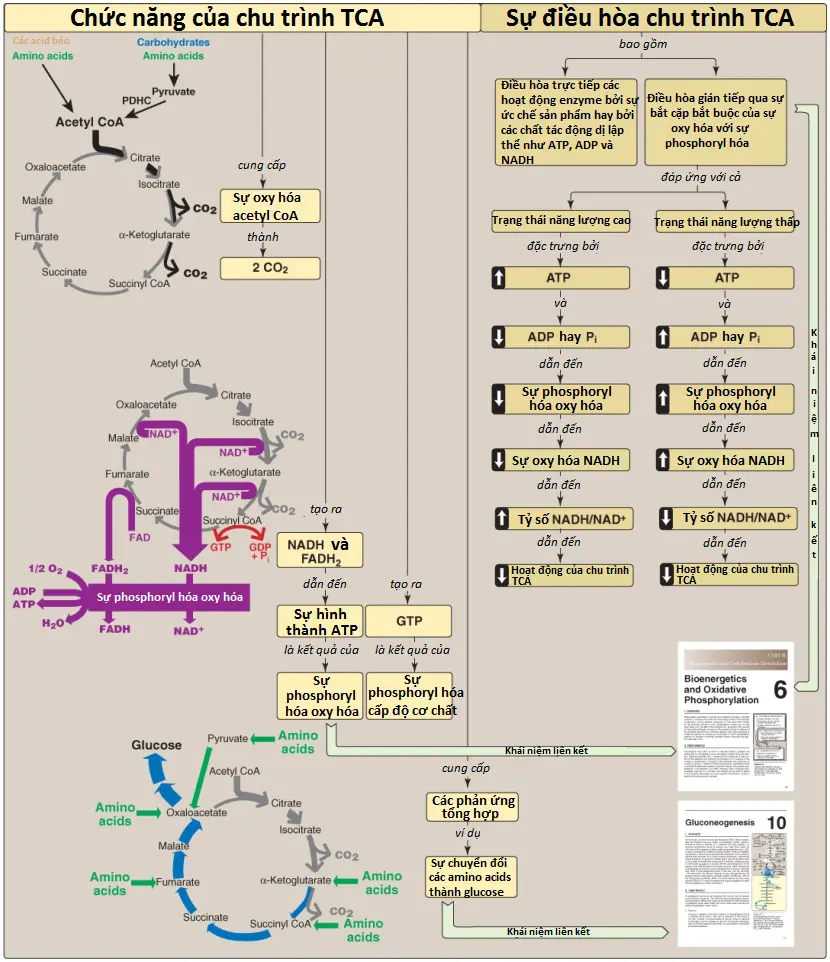
Chu trình tricarboxylic acid (tricarboxylic acid cycle) (chu trình TCA [TCA cycle]) cũng có thể được gọi là chu trình citric acid (citric acid cycle) hay chu trình Krebs (Krebs cycle) và đóng nhiều vai trò quan trọng trong chuyển hóa. Nó là con đường cuối cùng nơi mà quá trình dị hóa oxy hóa carbohydrates, amino acids và các acid béo gặp nhau, các khung các carbon của chúng sẽ được chuyển thành carbon dioxide (CO2), như được thể hiện trên Hình 1. Quá trình oxy hóa này cung cấp năng lượng cho sự sản xuất của phần lớn ATP ở hầu hết động vật, bao gồm con người. Bởi vì chu trình TCA xảy ra hoàn toàn trong ty thể nên nó có liên quan gần với chuỗi truyền electron (electron transport chain – ETC), thành phần mà sẽ oxy hóa các coenzymes khử nicotinamide adenine dinucleotide (NADH) và flavin adenine dinucleotide (FADH2) được tạo ra bởi chu trình. Chu trình TCA là một con đường hiếu khí bởi vì oxygen (O2) thì được cần đến như là chất nhận electron cuối cùng. Các phản ứng như sự dị hóa của một số amino acids tạo ra các trung gian của chu trình và được gọi là là các phản ứng bổ sung (anaplerotic reactions [“anaplerotic” trong tiếng Hy Lạp có nghĩa là “làm đầy”]). Chu trình TCA cũng cung cấp các trung gian cho một số phản ứng đồng hóa quan trọng, như sự hình thành glucose từ các khung carbon của một số amino acids và sự tổng hợp của một số amino acids (sẽ nói đến trong các bài viết sau) và heme (sẽ nói đến trong các bài viết sau). Vì thế, chu trình này không nên được xem là một hệ thống khép kín mà thay vào đó, nên xem nó như là một hệ thống mở với các hợp chất đi vào và đi ra cần thiết.
II. Các phản ứng của chu trình
Trong chu trình TCA, oxaloacetate (OAA) đầu tiên được ngưng tụ với một nhóm acetyl từ acetyl coenzyme A (CoA) và sau đó nó sẽ được tái tạo khi mà chu trình được hoàn thành (xem Hình 1). Hai nguyên tử carbon đi vào trong chu trình dưới dạng acetyl CoA và hai nguyên tử carbon rời khỏi chu trình dưới dạng CO2. Vì thế, sự đi vào trong một vòng chu trình TCA không dẫn đến sự hình thành hay tiêu thụ tổng quan của các trung gian.
A. Sự sản xuất acetyl CoA
Nguồn acetyl CoA chính cho chu trình TCA là từ quá trình decarboxyl hóa oxy hóa pyruvate bởi phức hợp đa enzyme pyruvate dehydrogenase (pyruvate dehydrogenase complex – [PDH complex hay PDHC]). Tuy nhiên, PDHC thì không phải là một thành phần của chu trình TCA. Pyruvate, sản phẩm cuối cùng của quá trình đường phân, được vận chuyển từ bào tương vào trong chất nền ty thể bởi chất mang pyruvate của màng trong ty thể. Trong chất nền ty thể, PDHC chuyển pyruvate thành acetyl CoA. (Chú ý: Sự oxy hóa acid béo là một nguồn acetyl CoA khác nữa [sẽ nói đến trong các bài viết sau]).
1. Các enzymes thành phần của PDHC: PDHC là một tập hợp protein gồm nhiều bản sao của 3 enzymes, pyruvate decarboxylase ([E1] đôi khi được gọi là PDH), dihydrolipoyl transacetylase (E2) và dihydrolipoyl dehydrogenase (E3). Mỗi trong số này hoạt hóa một phần của phản ứng tổng quan (Hình 2). Sự liên kết vật lý của chúng giúp kết nối các phản ứng theo một trình tự thích hợp mà không có sự giải phóng ra các trung gian. Ngoài các enzymes tham gia vào trong sự chuyển đổi pyruvate thành acetyl CoA, PDHC cũng gồm hai enzymes điều hòa, pyruvate dehydrogenase kinase (PDH kinase) và pyruvate dehydrogenase phosphatase (PDH phosphatase).
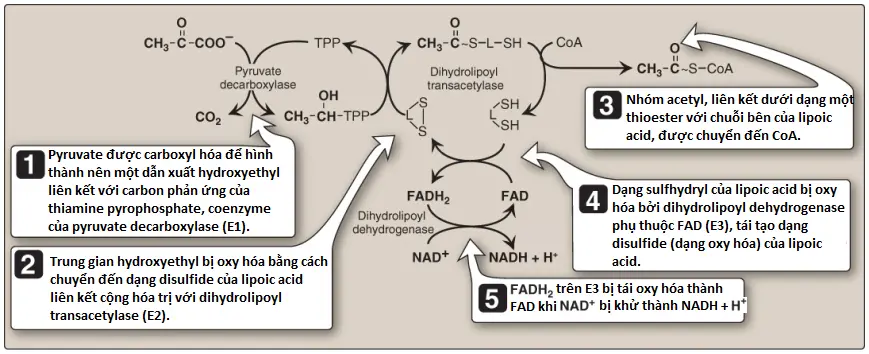
2. Coenzymes: PDHC chứa 5 coenzymes mà đóng vai trò như là các chất mang hay các chất oxy hóa đối với các trung gian của các phản ứng được thể hiện trong Hình 2. E1 cần thiamine pyrophosphate (TPP), E2 cần lipoic acid và CoA, và E3 cần FAD và NAD+. (Chú ý: TPP, lipoic acid và FAD được liên kết chặt chẽ với các enzymes và đóng vai trò như là các coenzymes – các nhóm phụ [prosthetic groups]).
Các sự suy giảm thiamine hay niacin có thể gây ra các vấn đề về hệ thống thần kinh trung ương nghiêm trọng. Điều này là bởi vì các tế bào não không thể tạo ra đủ ATP thông quan chu trình TCA nếu như PDHC bị bất hoạt. Wernicke – Korsakoff, một hội chứng bệnh não – loạn thần (encephalopathy-psychosis syndrome) do sự thiếu hụt thiamine, có thể phát hiện thấy ở những người có rối loạn sử dụng rượu.
3. Sự điều hòa: Các chỉnh sửa cộng hóa trị bởi 2 enzyme điều hòa của PDHC luân phiên hoạt hóa và bất hoạt E1. PDH kinase phosphoryl hóa và bất hoạt E1, ngược lại PDH phosphatase dephosphoryl hóa và hoạt hóa E1 (Hình 3). Chính kinase được hoạt hóa dị lập thể bởi ATP, acetyl CoA và NADH. Vì thế, trong sự có mặt của các sản phẩm cao năng lượng này, PDHC sẽ bị bất hoạt. (Chú ý: Thực sự là sự tăng lên trong các tỷ số ATP/ADP [adenosine diphosphate], NADH/NAD+ hay acetyl CoA/CoA sẽ ảnh hưởng đến hoạt động enzyme).
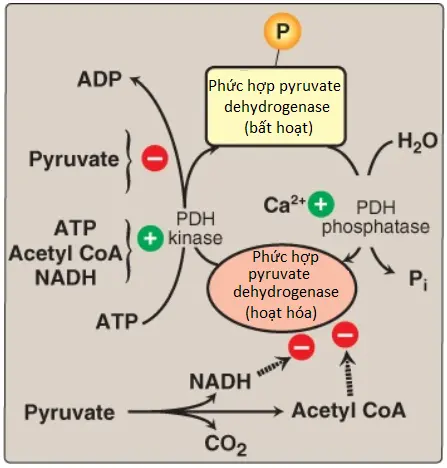
Pyruvate là một chất ức chế PDH kinase mạnh. Vì thế, nếu như các nồng độ pyruvate tăng lên thì E1 sẽ được hoạt hóa một cách tối đa. Canxi (Ca2+) là một chất kích hoạt mạnh của PDH phosphate, kích thích hoạt động của E1. Điều này thì cực kỳ quan trọng trong cơ xương, nơi mà Ca2+ được giải phóng trong suốt sự co cơ sẽ kích thích PDHC và vì thế, giúp sản xuất ra năng lượng. (Chú ý: Mặc dù sự điều hòa cộng hóa trị bởi kinase và phosphatase là chủ yếu nhưng PDHC cũng bị ảnh hưởng bởi sự ức chế sản phẩm (product inhibition) [NADH và acetyl CoA]).
4. Sự thiếu hụt: Một sự thiếu hụt các tiểu đơn vị α của thành phần tetramer E1 của PDHC, mặc dù rất hiếm nhưng nó là nguyên nhân hóa sinh thường gặp nhất của nhiễm toan lactic bẩm sinh (congenital lactic acidosis). Sự suy giảm có thể gây ra sự giảm khả năng chuyển pyruvate thành acetyl CoA, làm cho pyruvate được chuyển đổi thành lactate thông qua lactate dehydrogenase. Điều này gây ra các vấn đề nhất định cho não, nơi mà dựa vào chu trình TCA cho phần lớn năng lượng của nó và đặc biệt nhạy cảm với sự nhiễm toan. Các triệu chứng thì thay đổi và bao gồm thoái hóa thần kinh (neurodegeneration), co cứng cơ (muscle spasticity) và ở thể khởi sơ sinh (neonatal-onset form) là tử vong sớm. Gene của tiểu đơn vị α thì nằm trên nhiễm sắc thể X. Sự di truyền của chỉ một nhiễm sắc thể X đột biến sẽ tạo thành bệnh; kiểu di truyền này là kiểu trội liên kết nhiễm sắc thể X, với cả hai giới nam và nữ đều bị ảnh hưởng. Mặc dù không có phương pháp điều trị đã được xác định đối với vấn đề thiếu hụt PDHC nhưng sự hạn chế carbohydrate và sự bổ sung thiamine trong chế độ ăn có thể làm giảm các triệu chứng ở các bệnh nhân nhất định.
Hội chứng Leigh (Leigh syndrome) (bệnh não – tủy hoại tử bán cấp [subacute necrotizing encephalomyelopathy]) là một rối loạn thoái hóa thần kinh hiếm, tiến triển được gây ra bởi các khiếm khuyết trong sự sản xuất ATP của ty thể, chủ yếu là do các đột biến trong các genes mã hóa cho các proteins của PDHC, ETC hay ATP synthase. Cả DNA nhân và ty thể đều có thể bị ảnh hưởng.
5. Ngộ độc arsenic: Như đã được mô tả trong các bài viết trước, arsenic hóa trị 5 (arsenate) có thể can thiệp vào quá trình đường phân ở bước glyceraldehyde 3-phosphate, bằng cách đó, làm giảm sự sản xuất ATP. Tuy nhiên, ngộ độc arsenic chủ yếu là do sự ức chế của các phức hợp enzyme cần lipoic acid như là một coenzyme, bao gồm PDH, α-ketoglutarate dehydrogenase và α-keto acid phân nhánh dehydrogenase (branched-chain α-keto acid dehydrogenase) (sẽ bàn đến trong các bài viết sau nhé). Arsenite (dạng hóa trị 3 của arsenic) hình thành nên một phức hợp ổn định với các nhóm thiol (-SH) của lipoic acid, làm cho hợp chất đó không thể đóng vai trò như là một coenzyme. Khi nó liên kết với lipoic acid trong PDHC thì pyruvate (và tiếp theo là lactate) sẽ tích tụ lại. Cũng giống với sự suy giảm PDHC, tình trạng này sẽ ảnh hưởng đến não, gây ra các rối loạn thần kinh và tử vong.
B. Sự tổng hợp citrate
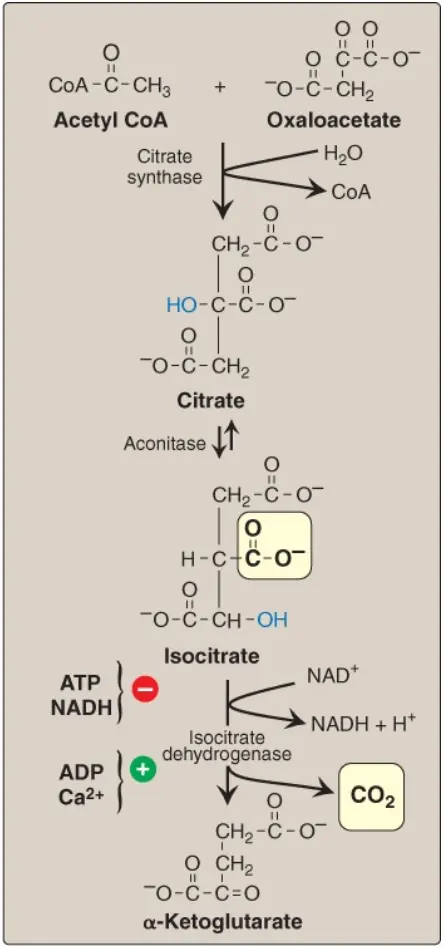
Sự ngưng tụ không thể đảo ngược của acetyl CoA và OAA để hình thành nên citrate (một TCA) được xúc tác bởi citrate synthase, enzyme khởi động của chu trình TCA (Hình 4). Sự ngưng tụ aldol này này có một sự chênh lệch âm lớn trong năng lượng tự do tiêu chuẩn ([ΔG0]), tạo điều kiện rất thuận lợi để hình thành citrate. Enzyme bị ức chế bởi citrate (ức chế sản phẩm). Sự có mặt của cơ chất là một phương tiện khác của sự điều hòa citrate synthase. Sự liên kết của OAA làm tăng mạnh ái tính của enzyme đối với acetyl CoA. (Chú ý: Citrate, ngoài là một trung gian trong chu trình TCA, nó còn là một nguồn acetyl CoA cho sự tổng hợp trong bào tương của các acid béo và cholesterol. Citrate cũng ức chế phosphofructokinase-1 [PFK-1], enzyme giới hạn tốc độ của quá trình đường phân và hoạt hóa acetyl CoA carboxylase [enzyme giới hạn tốc độ của sự tổng hợp acid béo, sẽ nói đến trong các bài viết sau nhé]).
C. Sự isomer hóa citrate
Citrate được isomer hóa thành isocitrate thông qua sự chuyển nhóm hydroxyl được xúc tác bởi aconitase (aconitate hydratase), một protein sắt – lưu huỳnh (xem Hình 4). (Chú ý: Aconitase bị ức chế bởi fluoroacetate, một độc tố thực vật được sử dụng như là một thuốc trừ sâu. Fluoroacetate được chuyển thành fluoroacetyl CoA, thành phần mà ngưng tụ với OAA để hình thành nên fluorocitrate, một chất ức chế mạnh của aconitase).
D. Sự decarboxyl hóa oxy hóa isocitrate
Isocitrate dehydrogenase xúc tác quá trình decarboxyl hóa oxy hóa isocitrate không thể đảo ngược thành α-ketoglutarate, tạo ra phân tử NADH đầu tiên trong 3 phân tử được tạo ra bởi chu trình và giải phóng ra CO2 đầu tiên (xem Hình 4). Đây là một trong những bước giới hạn tốc độ của chu trình TCA. Enzyme được hoạt hóa về mặt dị lập thể bởi ADP (một tín hiệu năng lượng thấp) và Ca2+ và bị ức chế bởi ATP và NADH, các mức chất này tăng lên khi tế bào có các sự tích trữ năng lượng dồi dào.
E. Sự decarboxyl hóa oxy hóa α-ketoglutarate
Sự chuyển đổi không thể đảo ngược của α-ketoglutarate thành succinyl CoA được xúc tác bởi phức hợp α-ketoglutarate dehydrogenase, một tập hợp protein gồm nhiều bản sao của 3 enzymes (Hình 5). Cơ chế của quá trình decarboxyl hóa oxy hóa này thì rất giống với cơ chế được sử dụng cho sự chuyển đổi của pyruvate thành acetyl CoA bởi PDHC. Phản ứng giải phóng ra phân tử CO2 thứ hai và tạo ra phân tử NADH thứ hai của chu trình. Các coenzymes cần thiết là TPP, lipoic acid, FAD, NAD+ và CoA. Mỗi trong số chúng thực hiện chức năng như là một phần của cơ chế xúc tác theo một cách tương tự với những gì được mô tả với PDHC. ΔG0 âm lớn của phản ứng tạo điều kiện thuận lợi cho sự hình thành của succinyl CoA, một thioester cao năng tương tự với acetyl CoA. Phức hợp α-ketoglutarate dehydrogenase bị ức chế bởi các sản phẩm của nó, NADH và succinyl CoA, và được hoạt hóa bởi Ca2+. Tuy nhiên, nó thì không được điều hòa bởi các phản ứng phosphoryl hóa/dephosphoryl hóa như được mô tả với PDHC. (Chú ý: α-ketoglutarate cũng được sản xuất bởi quá trình deamine hóa oxy hóa và quá trình chuyển amine hóa của amino acid glutamate).
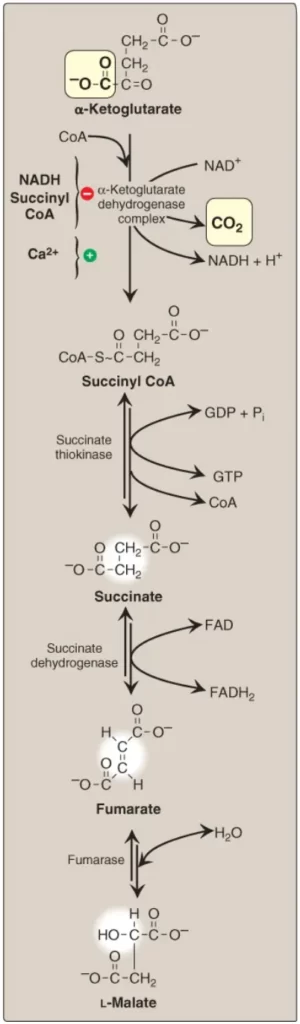
F. Sự phân cắt succinyl CoA
Succinate thiokinase (còn được gọi là succinyl CoA synthetase, được đặt tên theo phản ứng nghịch) phân cắt liên kết thioester cao năng lượng của succinyl CoA (xem Hình 5). Phản ứng này được bắt cặp với sự phosphoryl hóa của guanosine diphosphate (GDP) thành guanosine triphosphate (GTP). GTP và ATP thì có thể chuyển đổi qua lại một cách mạnh mẽ bởi phản ứng được xúc tác bởi nucleoside diphosphate kinase:
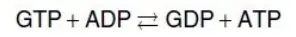
Sự tạo thành GTP bởi succinate thiokinase là một ví dụ khác của sự phosphoryl hóa cấp độ cơ chất (substrate-level phosphorylation). (Chú ý: Succinyl CoA cũng được sản xuất từ propionyl CoA có nguồn gốc từ sự chuyển hóa các acid béo có số nguyên tử carbon lẻ và từ sự chuyển hóa của một số amino acid. Nó có thể được chuyển thành pyruvate cho quá trình tân tạo đường (gluconeogenesis) [sẽ nói đến trong các bài viết tiếp theo] hoặc được sử dụng trong quá trình tổng hợp heme [sẽ nói đến trong các bài viết tiếp theo]).
G. Sự oxy hóa succinate
Succinate được oxy hóa thành fumarate bởi succinate dehydrogenase khi coenzyme FAD của nó được khử thành FADH2 (xem Hình 5). Succinate dehydrogenase là enzyme duy nhất của chu trình TCA mà vùi trong màng trong ty thể. Và do đó, nó đóng vai trò là Complex II trong chuỗi truyền electron (electron transport chain – ETC). (Chú ý: FAD, chứ không phải NAD+, là chất nhận electron bởi vì độ khử của succinate thì không đủ để khử NAD+).
H. Sự hydrate hóa fumarate
Fumarate được hydrate hóa thành malate trong một phản ứng thuận nghịch tự do được xúc tác bởi fumarase (fumarate hydratase, xem Hình 5). (Chú ý: Fumarate cũng được tạo ra bởi chu trình urea, trong sự tổng hợp purine và trong suốt quá trình dị hóa các amino acid phenylalanine và tyrosine).
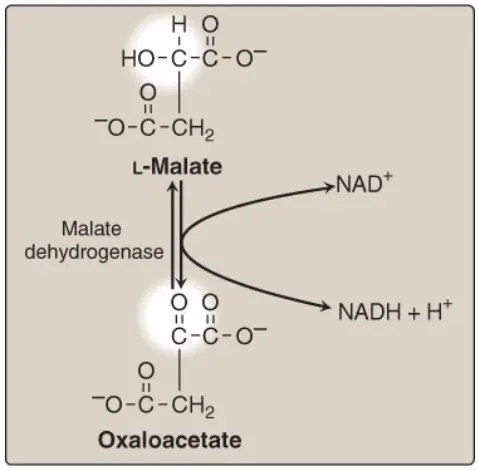
I. Sự oxy hóa malate
Malate được oxy hóa thành OAA bởi malate dehydrogenase (Hình 6). Phản ứng này tạo ra phân tử NADH thứ ba và cũng là cuối cùng của chu trình. ΔG0 của phản ứng là dương nhưng phản ứng được điều khiển theo hướng OAA bởi phản ứng tỏa năng lượng mạnh được xúc tác bởi citrate synthase. (Chú ý: OAA cũng được tạo ra bởi sự chuyển amine hóa của amino acid aspartic acid).
III. Năng lượng được sản xuất bởi chu trình
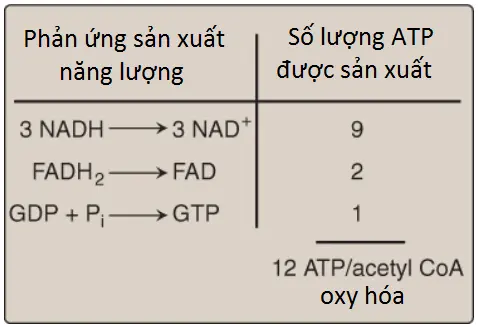
Bốn cặp electron được truyền đi trong suốt một vòng của chu trình TCA: 3 cặp đến từ sự khử 3 NAD+ thành NADH và một cặp đến từ sự khử FAD thành FADH2. Sự oxy hóa một phân tử NADH bởi chuỗi truyền electron (ETC) dẫn đến sự hình thành của 3 phân tử ATP. Tổng lượng phân tử ATP được tạo thành từ sự oxy hóa một phân tử acetyl CoA được thể hiện trên Hình 7. Hình 8 tóm tắt các phản ứng của chu trình TCA. (Chú ý: Chu trình không liên quan đến sự tổng hợp hay tiêu thụ toàn phần của các trung gian. Hai nguyên tử carbon đi vào chu trình dưới dạng acetyl CoA được cân bằng bởi hai phân tử CO2 đi ra khỏi chu trình).
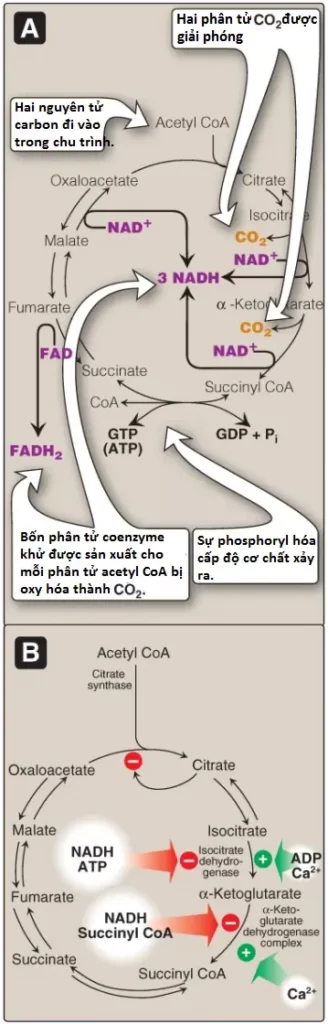
IV. Sự điều hòa chu trình
Ngược lại với quá trình đường phân, là một quá trình được điều hòa chủ yếu bởi PFK-1, chu trình TCA thì được kiểm soát bởi sự điều hòa của một số enzymes (xem Hình 8). Quan trọng nhất trong số các enzymes điều hòa này là các enzymes mà xúc tác cho các phản ứng có ΔG0 âm lớn: citrate synthase, isocitrate dehydrogenase và phức hợp α-ketoglutarate dehydrogenase. Các đương lượng khử cần cho sự phosphoryl hóa oxy hóa được tạo ra bởi PDHC và chu trình TCA, và cả hai quá trình đều được tăng cường điều hòa trong đáp ứng với một sự giảm trong tỷ số ATP/ADP.
V. Tổng hợp bài viết
- Trong chu trình TCA (TCA cycle), còn được gọi là chu trình Krebs (Krebs cycle), pyruvate bị decarboxyl hóa oxy hóa bởi PDHC, tạo ra acetyl CoA (Hình 9).
- Đa enzyme PDHC cần 5 coenzymes: TPP, lipoic acid, flavin adenine dinucleotide (FAD), nicotinamide adenine dinucleotide (NAD+) và CoA.
- PDHC được điều hòa bởi sự chỉnh sửa cộng hóa trị của E1, bởi PDH kinase và PDH phosphatase. Sự phosphoryl hóa sẽ ức chế E1.
- PDH kinase được hoạt hóa về mặt dị lập thể bởi ATP, acetyl CoA và NADH và bị ức chế bởi pyruvate. Phosphatase được hoạt hóa bởi canxi (Ca2+).
- Sự thiếu hụt pyruvate decarboxylase (pyruvate decarboxylase deficiency) là nguyên nhân sinh hóa thường gặp nhất của tình trạng nhiễm toan lactic bẩm sinh (congenital lactic acidosis). Não thì sẽ bị ảnh hưởng đặc biệt trong rối loạn trội liên kết nhiễm sắc thể X này (X-linked dominant disorder).
- Ngộ độc arsenic (arsenic poisoning) gây ra sự bất hoạt của PDHC bằng cách liên kết với lipoic acid. Trong chu trình TCA, citrate được tổng hợp từ OAA và acetyl CoA bởi citrate synthase, thành phần bị ức chế bởi sản phẩm.
- Citrate được isomer hóa thành isocitrate bởi aconitase (aconitate hydratase). Isocitrate bị decarboxyl hóa oxy hóa bởi isocitrate dehydrogenase thành α-ketoglutarate, tạo ra CO2 và NADH. Enzyme bị ức chế bởi ATP và NADH và được hoạt hóa bởi ADP và Ca2+.
- α-ketoglutarate bị decarboxyl hóa oxy hóa thành succinyl CoA bởi phức hợp α-ketoglutarate dehydrogenase, tạo ra CO2 và NADH. Enzyme này thì rất giống với PDHC và sử dụng cùng các coenzymes.
- Phức hợp α-ketoglutarate dehydrogenase được hoạt hóa bởi Ca2+ và bị ức chế bởi NADH và succinyl CoA nhưng không được điều hòa về mặt cộng hóa trị. Succinyl CoA được phân tách bởi succinate thiokinase, tạo ra succinate và GTP. Đây là một ví dụ của sự phosphoryl hóa cấp độ cơ chất (substrate-level phosphorylation).
- Succinate bị oxy hóa thành fumarate bởi succinate dehydrogenase, tạo thành FADH2. Fumarate bị hydrate hóa thành malate bởi fumarase (fumarate hydratase) và malate bị oxy hóa thành OAA bởi malate dehydrogenase, tạo ra NADH.
- Ba NADH và một FADH2 được tạo ra bởi một vòng chu trình TCA.
- Sự tạo thành acetyl CoA bởi sự oxy hóa pyruvate thông qua PDHC cũng tạo ra một NADH. Sự oxy hóa NADH và FADH2 bởi chuỗi truyền electron (ETC) tạo ra 14 ATP. Nhóm phosphate tận cùng của GTP được tạo ra bởi sự phosphoryl hóa cấp độ cơ chất trong chu trình TCA có thể được chuyển đến ADP bởi nucleoside diphosphate kinase, tạo ra một ATP nữa.
- Vì thế, tổng cộng có 15 ATP được tạo ra từ sự oxy hóa pyruvate ty thể hoàn toàn thành CO2.
Các bạn có thể xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn có thể xem bài viết trước tại đây: https://docsachxyz.com/gioi-thieu-ve-chuyen-hoa-va-duong-phan-phan-2/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!







{kind=link}