





























I. Tổng quan
Một nguồn glucose máu hằng định là một yêu cầu tuyệt đối cho sự sống con người. Glucose là nguồn năng lượng rất được ưu tiên đối với não và là nguồn năng lượng cần thiết cho các tế bào mà có ít hoặc không có ty thể như các tế bào hồng cầu trưởng thành. Glucose cũng là một nguồn năng lượng thiết yếu cho cơ hoạt động thể thao, nơi mà nó là cơ chất cho quá trình đường phân kỵ khí (anaerobic glycolysis). Glucose máu có thể thu được từ 3 nguồn chính: chế độ ăn, thoái hóa glycogen và quá trình tân tạo đường (gluconeogenesis). Glucose và các tiền thân glucose ăn vào trong chế độ ăn như tinh bột (một polysaccharide), disaccharides và monosaccharides, là không thường xuyên, phụ thuộc vào chế độ ăn, không phải lúc nào cũng là một nguồn glucose máu bảo đảm. Ngược lại, quá trình tân tạo đường có thể cung cấp sự tổng hợp glucose duy trì nhưng nó thì sẽ hơi chậm trong đáp ứng với một mức glucose máu đang hạ xuống. Vì thế, cơ thể sẽ phát triển các cơ chế tích trữ một nguồn glucose ở dạng có thể huy động nhanh, được gọi là glycogen. Trong trường hợp không có nguồn glucose trong chế độ ăn, đường này sẽ được giải phóng một cách nhanh chóng vào trong máu từ glycogen của gan. Tương tự, glycogen cơ sẽ được thoái hóa một cách rộng rãi trong cơ đang luyện tập thể thao để cung cấp cho mô này một nguồn năng lượng quan trọng. Khi các sự tích trữ glycogen cạn kiệt, các mô chuyên biệt sẽ tổng hợp glucose mới, sử dụng glycerol, lactate, pyruvate và các amino acids như là các nguồn carbon cho quá trình tân tạo đường (xem bài viết trước của mình). Hình 1 cho thấy các phản ứng của sự tổng hợp và thoái hóa glycogen như là một phần của các con đường chuyển hóa năng lượng thiết yếu.

II. Cấu trúc và chức năng
Các sự tích trữ glycogen chính được tìm thấy trong cơ xương và gan mặc dù hầu hết các tế bào khác tích trữ các lượng nhỏ glycogen cho nhu cầu sử dụng của chính chúng. Chức năng của glycogen cơ thì đóng vai trò như là một sự tích trữ nguyên liệu cho sự tổng hợp ATP trong suốt quá trình co cơ, trong khi mục đích của glycogen gan là để duy trì nồng độ glucose máu, đặc biệt là trong suốt các giai đoạn đầu của một quá trình nhịn ăn (Hình 2) (Chú ý: Glycogen gan có thể duy trì glucose máu trong dưới 24 giờ).

A. Các lượng glycogen trong gan và cơ
Gần 400 grams glycogen tạo thành 1% đến 2% trọng lượng tươi của khối cơ đang nghỉ ngơi, và khoảng 100 grams glycogen tạo thành đến 10% trọng lượng tươi của gan người trưởng thành khi ăn no. Điều gì giới hạn sự sản xuất glycogen ở các mức này thì vẫn chưa rõ ràng. Tuy nhiên, trong một sự bệnh tích trữ glycogen (glycogen storage diseases – GSDs) (xem Hình 8), lượng glycogen trong gan và/hoặc trong cơ có thể cao hơn đáng kể. (Chú ý: Trong cơ thể, khối lượng cơ thì lớn hơn nhiều so với khối lượng gan. Do đó, hầu hết glycogen của cơ thể thì được tìm thấy trong cơ xương).
B. Cấu trúc
Glycogen là một polysaccharide phân nhánh được tạo thành hoàn toàn từ α-D-glucose. Liên kết glycoside chính là một liên kết α(1→4). Sau trung bình khoảng 8 đến 14 gốc glucosyl thì sẽ có một nhánh chứa một liên kết α(1→6) (Hình 3). Một phân tử glycogen đơn có thể chứa đến 55,000 gốc glucosyl. Các polymers glucose này tồn tại dưới dạng các hạt bào tương hình cầu lớn mà cũng chứa hầu hết các enzymes cần cho sự tổng hợp và thoái hóa glycogen.

C. Biến động tích trữ glycogen
Các tích trữ glycogen tăng lên trong suốt trạng thái ăn no và cạn kiệt trong suốt quá trình nhịn ăn. Glycogen cơ thì không bị ảnh hưởng bởi các giai đoạn nhịn ăn ngắn (một vài ngày) và chỉ giảm một cách trung bình trong giai đoạn nhịn ăn kéo dài (vài tuần). Glycogen cơ được tổng hợp để làm đầy lại các sự tích trữ trong cơ sau khi chúng cạn kiệt sau hoạt động thể thao mạnh. (Chú ý: Sự tổng hợp và thoái hóa glycogen tiếp diễn liên tục. Sự chênh lệch về tốc độ của hai quá trình này giúp xác định các mức glycogen tích trữ trong suốt các trạng thái sinh lý chuyên biệt).
III. Sự tổng hợp (quá trình tổng hợp glycogen)
Glycogen được tổng hợp từ các phân tử α-D-glucose. Quá trình xảy ra trong bào tương và cần năng lượng được cung cấp bởi ATP (cho sự phosphoryl hóa của glucose) và uridine triphosphate (UTP).
A. Sự tổng hợp uridine diphosphate glucose
α-D-glucose nối với uridine diphosphate (UDP) là nguồn gốc của tất cả các gốc glucosyl mà được thêm vào trong phân tử glycogen đang phát triển. UDP-glucose (Hình 4) được tổng hợp từ glucose 1-phosphate và UTP bởi UDP – glucose pyrophosphorylase (Hình 5). Pyrophosphate (PPi), sản phẩm thứ hai của phản ứng, được thủy phân thành 2 phosphate vô cơ (Pi) bởi pyrophosphatase. Quá trình thủy phân là quá trình tỏa năng lượng, điều mà đảm bảo rằng phản ứng xúc tác bởi UDP-glucose pyrophosphorylase diễn ra theo hướng của sự sản xuất ra UDP-glucose. (Chú ý: Glucose 1-phosphate được tạo ra từ glucose 6-phosphate bởi phosphoglucomutase. Glucose 1,6-bisphosphate là một trung gian bắt buộc trong phản ứng có thể đảo ngược này [Hình 6]).



B. Yêu cầu và sự tổng hợp đoạn mồi (primer)
Glycogen synthase xúc tác cho các liên kết α(1→4) trong glycogen. Enzyme này không thể khởi động sự tổng hợp chuỗi nhờ sử dụng glucose tự do như là một chất nhận của một phân tử glucose từ UDP-glucose. Thay vào đó, nó chỉ có thể kéo dài các chuỗi glucose đã tồn tại sẵn và vì thế, cần một đoạn mồi (primer). Một đoạn glycogen có thể đóng vai trò như là một đoạn mồi. Trong trường hợp không có một đoạn mồi thì dimer protein glycogenin có thể đóng vai trò như là một chất nhận glucose từ UDP-glucose (xem Hình 5). Nhóm hydroxyl chuỗi bên của tyrosine-194 trong protein là vị trí mà đơn vị glucosyl ban đầu sẽ nối vào. Bởi vì phản ứng được xúc tác bởi chính glycogenin thông qua sự tự glucosyl hóa nên glycogenin là một enzyme. Glycogenin sau đó xúc tác sự chuyển của ít nhất 4 phân tử glucose từ UDP-glucose, tạo ra một chuỗi glucosyl liên kết α(1→4) ngắn. Chuỗi ngắn này đóng vai trò như là một đoạn mồi mà có thể được kéo dài bởi glycogen synthase, enzyme được huy động bởi glycogenin. (Chú ý: Glycogenin liên quan với và hình thành nên lõi của hạt glycogen).
C. Sự kéo dài bởi glycogen synthase
Sự kéo dài của chuỗi glycogen liên quan với sự chuyển glucose từ UDP-glucose đến đầu không khử (nonreducing end) của chuỗi đang phát triển, hình thành nên một liên kết glycoside mới giữa nhóm hydroxyl anomeric của carbon thứ nhất của glucose hoạt hóa và carbon thứ tư của gốc glucosyl nhận (xem Hình 5). (Chú ý: Đầu không khử của một chuỗi carbohydrate là đầu mà trong đó carbon anomeric của phân tử đường cuối cùng được liên kết với một phân tử khác bởi một liên kết glycoside, làm cho phân tử đường cuối cùng không có tính khử). Enzyme chịu trách nhiệm cho việc hình thành nên các liên kết α(1→4) trong glycogen là glycogen synthase. (Chú ý: UDP được giải phóng khi liên kết glycoside α[1→4] được tạo thành có thể được phosphoryl hóa thành UTP bởi nucleoside diphosphate kinase [UDP + ATP ⇄ UTP + ADP]).
D. Sự hình thành nhánh
Nếu như không có enzyme tổng hợp khác tác động lên chuỗi thì cấu trúc hình thành sẽ là một chuỗi thẳng (không phân nhánh) của các gốc glucosyl được nối bởi các liên kết α(1→4). Một hợp chất như thế thì được tìm thấy trong các mô thực vật và được gọi là amylose. Ngược lại, glycogen có các nhánh trung bình nằm cách nhau 8 gốc glucosyl, tạo thành một cấu trúc dạng cây phân nhánh mạnh (xem Hình 3) mà dễ tan hơn so với amylose không phân nhánh rất nhiều. Sự phân nhánh cũng làm tăng số lượng các đầu không khử mà các gốc glucosyl mới có thể nối vào (và cũng từ đây các gốc này có thể bị loại bỏ như được mô tả trong phần tiếp theo của bài viết), bằng cách đó, làm tăng mạnh tốc độ mà sự tổng hợp glycogen có thể xảy ra và tăng nhanh kích thước của phân tử glycogen.
1. Sự tổng hợp nhánh: Các nhánh được tạo thành bởi hoạt động của enzyme phân nhánh (branching enzyme), amylo-α(1→4)→α(1→6)-transglycosylase. Enzyme này loại bỏ một tập hợp 6 đến 8 gốc glucosyl từ đầu không khử của chuỗi glycogen, bẻ gãy một liên kết α(1→4) trên chuỗi và nối nó với một gốc glucosyl không phải tận cùng bởi một liên kết α(1→6), vì thế, thực hiện chức năng như là một 4:6 transferase. Đầu tận không khử mới (xem “i” trong Hình 5), cũng như là đầu tận không khử cũ mà từ đó 6 đến 8 gốc bị loại bỏ (xem “o” trong Hình 5), bây giờ có thể kéo dài hơn bởi glycogen synthase.
2. Sự tổng hợp thêm nhánh: Sau khi việc kéo dài 2 đầu tận này được hoàn thành, 6 đến 8 gốc glucosyl tận của chúng có thể được loại bỏ và được sử dụng để tạo ra các nhánh khác.
IV. Sự thoái hóa (quá trình phân giải glycogen)
Con đường thoái hóa mà huy động glycogen tích trữ trong gan và các cơ xương thì không phải là sự đảo ngược của các phản ứng tổng hợp. Thay vào đó, một tập hợp các enzymes bào tương khác thì sẽ được sử dụng. Khi glycogen bị thoái hóa, sản phẩm chủ yếu là glucose 1-phosphate, thu được bởi sự bẻ gãy các liên kết glycoside α(1→4). Ngoài ra, glucose tự do được giải phóng từ mỗi gốc glucosyl có liên kết α(1→6) (điểm nhánh).
A. Làm ngắn chuỗi
Glycogen phosphorylase lần lượt cắt đứt các liên kết glycoside α(1→4) giữa các gốc glucosyl ở các đầu không khử của các chuỗi glycogen bởi quá trình phosphoryl phân (phosphorolysis) (tạo ra glucose 1-phosphate) cho đến khi 4 đơn vị glucosyl còn lại trên mỗi chuỗi ở một điểm nhánh (Hình 7). Cấu trúc còn lại được gọi là một dextrin giới hạn (limit dextrin) và phosphorylase không thể thoái hóa nó nữa (Hình 8). (Chú ý: Phosphorylase cần pyridoxal phosphate [một dẫn xuất của vitamin B6] đóng vai trò như là một coenzyme).

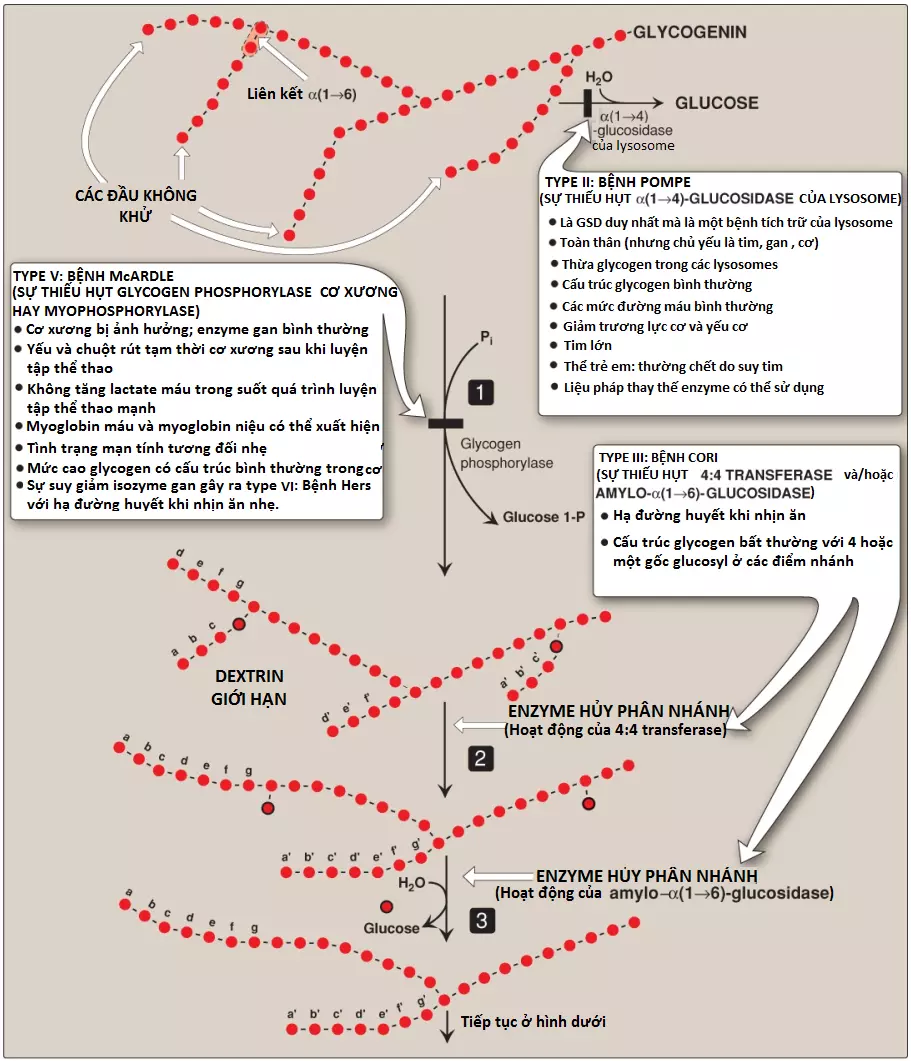
B. Sự loại bỏ nhánh
Các nhánh được loại bỏ bởi các hoạt động enzyme của một protein đơn hai chức năng (single bifunctional protein) là enzyme hủy phân nhánh (debranching enzyme) (xem Hình 8). Đầu tiên, hoạt động của oligo-α(1→4)→α(1→4)-glucantransferase giúp loại bỏ 3 trong 4 gốc glucosyl bên ngoài ở một nhánh. Tiếp theo, nó chuyển chúng đến đầu không khử của một chuỗi khác, do đó, sẽ làm dài chuỗi này. Vì thế, một liên kết α(1→4) bị bẻ gãy và một liên kết α(1→4) được hình thành và enzyme thực hiện chức năng như là một 4:4 transferase. Tiếp theo, gốc glucose còn lại được kết nối bởi liên kết α(1→6) thì sẽ được loại bỏ về mặt thủy phân bởi hoạt động của amylo-α(1→6)-glucosidase, giải phóng ra glucose tự do (không được phosphoryl hóa). Chuỗi glucosyl bây giờ sẽ sẵn sàng cho sự thoái hóa lại lần nữa bởi glycogen phosphorylase cho đến khi 4 đơn vị glucosyl trong nhánh tiếp theo được đạt đến.
C. Sự isomer hóa glucose 1-phosphate thành glucose 6-phosphate
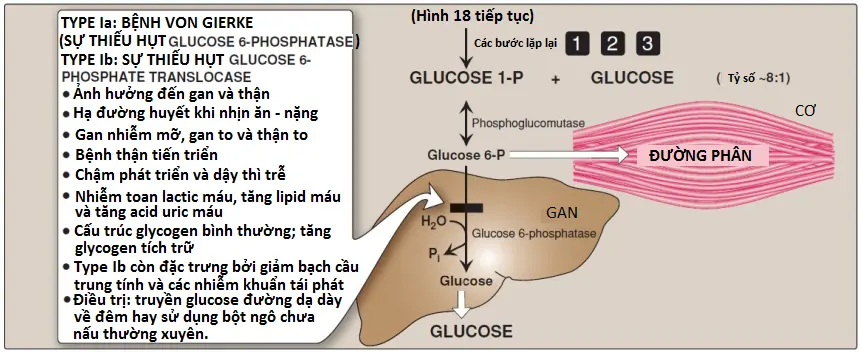
Glucose 1-phosphate, được sản xuất bởi glycogen phosphorylase, được isomer hóa trong bào tương thành glucose 6-phosphate bởi phosphoglucomutase (xem Hình 6). Trong gan, glucose 6-phosphate được vận chuyển vào trong lưới nội chất (ER) bởi glucose 6-phosphate translocase. Ở đó, nó được dephosphoryl hóa thành glucose bởi glucose 6-phosphatase (cùng enzyme được sử dụng trong bước cuối cùng của quá trình tân tạo đường, xem bài viết). Glucose sau đó được vận chuyển từ ER vào trong máu để giúp duy trì các mức glucose máu cho đến khi con đường tân tạo đường sản xuất glucose một cách tích cực. (Chú ý: Cơ thiếu glucose 6-phosphatase. Do đó, glucose 6-phosphate không thể được dephosphoryl hóa và đưa vào trong máu. Thay vào đó, nó đi vào trong quá trình đường phân, cung cấp năng lượng cần cho sự co cơ).
D. Sự thoái hóa glycogen của lysosome
Một lượng nhỏ (1% đến 3%) glycogen được thoái hóa bởi enzyme lysosome là acid α(1→4)-glucosidase (acid maltase). Mục đích của con đường tự thực này thì vẫn chưa được phát hiện. Tuy nhiên, một sự suy giảm của enzyme này sẽ gây ra sự tích tụ của glycogen trong các không bào trong các lysosomes, gây ra bệnh tích trữ glycogen type II (GSD type II) nghiêm trọng: bệnh Pompe (Pompe disease) (xem Bảng 1 và Hình 8). (Chú ý: Bệnh Pompe, gây ra bởi sự suy giảm acid maltase, là bệnh tích trữ glycogen duy nhất mà là một bệnh tích trữ của lysosome).
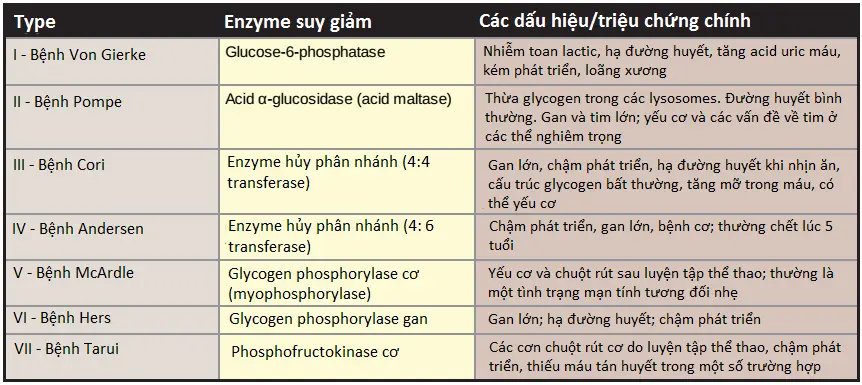
Các bệnh tích trữ lysosome (lysosomal storage diseases) là các rối loạn di truyền được đặc trưng bởi sự tích tụ các lượng carbohydrates hay lipids bất thường chủ yếu do sự giảm thoái hóa của lysosome do sự vắng mặt hoặc giảm hoạt động hay giảm lượng acid hydrolase đặc hiệu của lysosome mà bình thường sẽ chịu trách nhiệm cho sự thoái hóa của nó.
V. Sự điều hòa tổng hợp glycogen và phân giải glycogen
Bởi vì tầm quan trọng của việc duy trì các mức glucose máu nên sự tổng hợp và thoái hóa dạng tích trữ glycogen thì được điều hòa một cách chặt chẽ. Trong gan, sự tổng hợp glycogen sẽ tăng cường trong suốt các thời kỳ khi mà cơ thể được ăn đầy đủ, ngược lại quá trình phân giải glycogen sẽ tăng cường trong suốt các thời kỳ nhịn ăn. Trong cơ xương, quá trình phân giải glycogen xảy ra trong suốt quá trình luyện tập thể thao tích cực và quá trình tổng hợp glycogen sẽ bắt đầu sớm khi cơ được nghỉ ngơi trở lại.
Sự điều hòa tổng hợp và thoái hóa được thực hiện trên hai mức độ. Đầu tiên, glycogen synthase và glycogen phosphorylase được điều hòa về mặt hormone (bởi sự phosphoryl hóa/dephosphoryl hóa cộng hóa trị) để đáp ứng các nhu cầu của cơ thể nói chung. Thứ hai, cùng các enzymes này sẽ được điều hòa dị lập thể (bởi các phân tử tác động [effector molecules]) để đáp ứng các nhu cầu của mô nhất định.
A. Sự hoạt hóa cộng hóa trị của quá trình tổng hợp glycogen
Sự liên kết của các hormones như glucagon hay epinephrine với các thụ cảm thể bắt cặp G protein của màng tế bào sẽ truyền các tín hiệu về nhu cầu đối với việc glycogen cần được thoái hóa, hoặc về các mức glucose máu tăng lên khi cơ đang luyện tập thể thao.
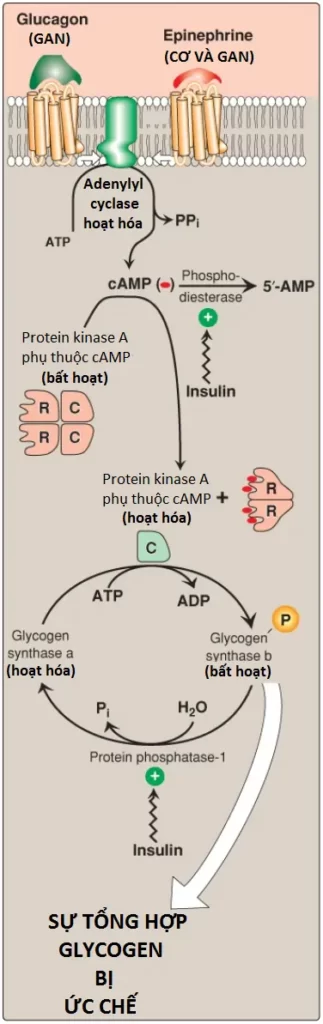
1. Sự hoạt hóa protein kinase A: Sự liên kết của glucagon hay epinephrine với GPCR (G protein-coupled receptors [các thụ cảm thể bắt cặp protein G]) tế bào gan đặc hiệu hay của epinephrine với GPCR tế bào cơ đặc hiệu sẽ gây ra sự hoạt hóa qua trung gian G protein của adenylyl cyclase. Enzyme này xúc tác sự tổng hợp của adenosine monophosphate vòng (cyclic adenosine monophosphate – cAMP), chất mà sẽ hoạt hóa protein kinase A phụ thuộc cAMP (PKA). cAMP liên kết với hai tiểu đơn vị điều hòa của tetramer PKA, giải phóng 2 tiểu đơn vị xúc tác hoạt hóa (Hình 9). PKA sau đó phosphoryl hóa một vài enzymes của chuyển hóa glycogen, ảnh hưởng đến hoạt động của chúng. (Chú ý: Khi cAMP bị loại bỏ, tetramer PKA bất hoạt sẽ hồi dạng).

2. Sự hoạt hóa phosphorylase kinase: Phosphorylase kinase tồn tại ở hai dạng: một dạng “b” bất hoạt và một dạng “a” hoạt hóa. PKA hoạt hóa phosphoryl hóa dạng bất hoạt “b” của phosphorylase kinase, tạo thành dạng “a” hoạt hóa (xem Hình 9).
3. Sự hoạt hóa glycogen phosphorylase: Glycogen phosphorylase cũng tồn tại trong một dạng “b” bất hoạt dephosphoryl hóa và một dạng “a” hoạt hóa phosphoryl hóa. Phosphorylase kinase a là enzyme duy nhất mà phosphoryl hóa glycogen phosphorylase b thành dạng “a” hoạt hóa của nó, thành phần mà sau đó sẽ bắt đầu quá trình phân giải glycogen (xem Hình 9).
4. Sự khuếch đại tín hiệu: Trình tự các phản ứng được mô tả ở trên sẽ hoạt hóa quá trình phân giải glycogen (glycogenolysis). Số lượng lớn các bước tuần tự đóng vai trò trong việc khuếch đại tác động của tín hiệu hormone, nghĩa là, một số ít phân tử hormone liên kết với GPCR của chúng làm cho một số phân tử PKA được hoạt hóa mà mỗi phân tử này có thể hoạt hóa nhiều phân tử phosphorylase kinase. Điều này dẫn đến sự sản xuất của nhiều phân tử glycogen phosphorylase a hoạt hóa mà có thể thoái hóa glycogen.
5. Sự duy trì trạng thái phosphoryl hóa: Các nhóm phosphate được thêm vào trong phosphorylase kinase và phosphorylase trong đáp ứng với cAMP thì được duy trì bởi vì enzyme mà loại bỏ nhóm phosphate về mặt thủy phân, protein phosphatase-1 (PP1), thì bị bất hoạt bởi các proteins ức chế mà cũng được phosphoryl hóa và hoạt hóa trong đáp ứng với cAMP (xem Hình 9). Bởi vì insulin cũng hoạt hóa phosphodiesterase, enzyme mà giúp thoái hóa cAMP nên nó cũng đối kháng lại các tác động của glucagon và epinephrine.
B. Sự ức chế cộng hóa trị của quá trình tổng hợp glycogen
Enzyme điều hòa trong quá trình tổng hợp glycogen (glycogenesis), glycogen synthase, cũng tồn tại ở hai dạng, dạng “a” hoạt hóa và dạng “b” bất hoạt. Tuy nhiên, ngược lại với phosphorylase kinase và phosphorylase, dạng hoạt hóa của glycogen synthase là khi được dephosphoryl hóa, ngược lại, dạng bất hoạt là khi bị phosphoryl hóa ở một vài vị trí trên enzyme, với mức độ bất hoạt tỷ lệ với mức độ phosphoryl hóa (Hình 10). Sự phosphoryl hóa được xúc tác bởi một vài protein kinases khác nhau trong đáp ứng với cAMP (ví dụ, PKA và phosphorylase kinase) hay các cơ chế truyền tín hiệu khác. Glycogen synthase b có thể được chuyển lại thành dạng “a” bởi PP1. Hình 11 tổng hợp sự điều hòa cộng hóa trị của chuyển hóa glycogen.

C. Sự điều hòa dị lập thể của quá trình tổng hợp glycogen và quá trình phân giải glycogen
Ngoài các tín hiệu hormone, glycogen synthase và glycogen phosphorylase còn đáp ứng với các mức của các chất chuyển hóa và các nhu cầu năng lượng của tế bào. Quá trình tổng hợp glycogen được kích thích khi mà các mức glucose và năng lượng cao, ngược lại quá trình phân giải glycogen thì tăng lên khi các mức glucose và năng lượng thấp. Sự điều hòa dị lập thể này cho phép một sự đáp ứng nhanh với các nhu cầu của một tế bào và có thể khống chế các tác động của sự điều hòa cộng hóa trị qua trung gian hormone. (Chú ý: Các dạng “a” và “b” của các enzymes dị lập thể của quá trình chuyển hóa glycogen thì ở trạng thái cân bằng giữa cấu hình R [thư giãn (relaxed), hoạt động hơn] và cấu hình T [căng thẳng (tense), ít hoạt động hơn]). Sự liên kết của các chất tác động sẽ dịch chuyển cân bằng và thay đổi hoạt động enzyme mà không làm thay đổi một cách trực tiếp sự chỉnh sửa cộng hóa trị).
1. Sự điều hòa trong trạng thái ăn no: Trong trạng thái ăn no, glycogen synthase b trong cả gan và cơ thì được hoạt hóa về mặt dị lập thể bởi glucose 6-phosphate, là chất mà xuất hiện với các nồng độ tăng lên (Hình 12). Ngược lại, glycogen phosphorylase a bị ức chế về mặt dị lập thể bởi glucose 6-phosphate, cũng như là bởi ATP, một tín hiệu cao năng lượng. Chú ý rằng trong gan, nhưng không có trong cơ, glucose tự do cũng là một chất ức chế dị lập thể của glycogen phosphorylase a.
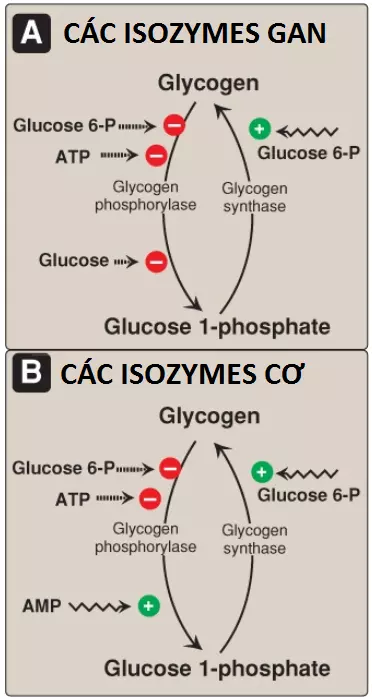
2. Sự hoạt hóa quá trình tổng hợp glycogen bởi AMP: Glycogen phosphorylase cơ (myophosphorylase) mà không có isozyme trong gan, thì sẽ hoạt hóa trong sự có mặt của các nồng độ AMP cao, xảy ra dưới các tình trạng cực kỳ thiếu oxygen mô và cạn kiệt ATP. AMP liên kết với glycogen phosphorylase b, gây ra sự hoạt hóa của nó mà không có sự phosphoryl hóa xảy ra (xem Hình 9). Nhớ lại rằng là AMP cũng hoạt hóa phosphofructokinase-1 của quá trình đường phân, cho phép glucose từ quá trình phân giải glycogen được oxy hóa.
3. Sự hoạt hóa quá trình phân giải glycogen bởi canxi: Canxi (Ca2+) được giải phóng vào trong cơ tương của các tế bào cơ trong sự đáp ứng với kích thích thần kinh và trong gan trong sự đáp ứng với việc epinephrine liên kết với các thụ cảm thể α1-adrenergic. Ca2+ liên kết với calmodulin (CaM), thành phần được phân bố rộng rãi nhất của một họ các proteins nhỏ liên kết với Ca2+. Sự liên kết của 4 phân tử Ca2+ với CaM sẽ khởi động một sự thay đổi cấu hình sao cho phức hợp Ca2+-CaM hoạt hóa liên kết và hoạt hóa các phân tử protein, thường là các enzymes mà bất hoạt khi không có mặt phức hợp này (Hình 13). Vì thế, CaM thực hiện chức năng như là một tiểu đơn vị thiết yếu của nhiều phức hợp protein. Một protein như thế chính là tetramer phosphorylase kinase, thành phần mà dạng “b” của nó được hoạt hóa bởi sự liên kết của Ca2+ với tiểu đơn vị δ của nó (CaM) mà không cần kinase được phosphoryl hóa bởi PKA. (Chú ý: Epinephrine ở các thụ cảm thể β-adrenergic truyền tín hiệu thông quan một sự tăng lên trong cAMP, không phải thông qua sự tăng lên trong Ca2+).
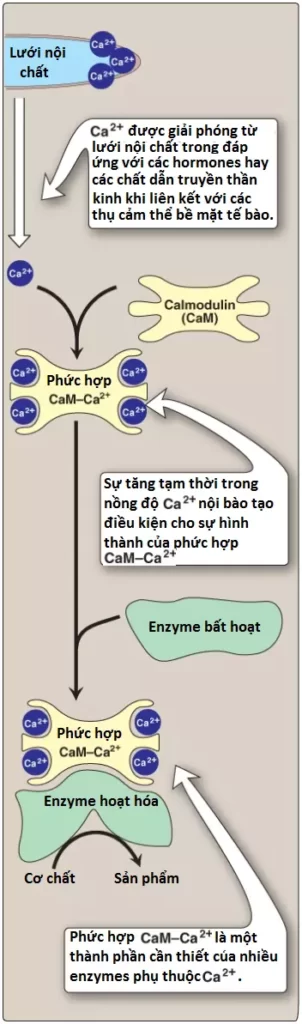
a. Sự hoạt hóa phosphorylase kinase cơ: Trong suốt quá trình co cơ, có một nhu cầu nhanh và khẩn cấp đối với ATP. Nó được cung cấp bởi sự thoái hóa của glycogen cơ thành glucose 6-phosphate, thành phần mà đi vào quá trình đường phân. Các xung động thần kinh gây ra sự khử cực của màng tế bào, điều này sẽ thúc đẩy sự giải phóng Ca2+ từ lưới cơ tương vào trong cơ tương của tế bào cơ. Ca2+ liên kết với tiểu đơn vị CaM và phức hợp này sẽ hoạt hóa phosphorylase kinase b của cơ (xem Hình 9).
b. Sự hoạt hóa phosphorylase kinase của gan: Trong suốt quá trình stress sinh lý, epinephrine được giải phóng từ tủy thượng thận và truyền tín hiệu về nhu cầu đối với glucose máu. Glucose này ban đầu đến từ quá trình phân giải glycogen của gan. Sự liên kết của epinephrine với α1-adrenergic GPCR của tế bào gan sẽ hoạt hóa một trình tự các phản ứng phụ thuộc phospholipid mà gây ra sự di chuyển của Ca2+ từ ER vào trong bào tương. Một phức hợp Ca2+-CaM hình thành và hoạt hóa phosphorylase kinase b của gan. Chú ý rằng Ca2+ giải phóng cũng giúp hoạt hóa protein kinase C, thành phần mà có thể phosphoryl hóa và bất hoạt glycogen synthase a.
VI. Các bệnh tích trữ glycogen (GSDs)
GSDs (glycogen storage diseases) là một nhóm các bệnh di truyền được gây ra bởi các khiếm khuyết trong các enzymes mà cần cho sự thoái hóa glycogen (glycogen degradation) hay hiếm gặp hơn là sự tổng hợp glycogen (glycogen synthesis). Các triệu chứng thường gặp nhất là hạ đường huyết (hypoglycemia) (glucose máu thấp), gan to, phát triển chậm và yếu cơ hay chuột rút cơ.
Các rối loạn này gây sự hình thành của glycogen mà có một cấu trúc bất thường hoặc gây ra sự tích tụ của các lượng glycogen bình thường quá mức trong các mô đặc hiệu như là kết quả của sự suy giảm quá trình thoái hóa. Một enzyme đặc hiệu có thể bị khiếm khuyết trong một mô duy nhất, như gan (gây ra sự hạ đường huyết) hay cơ (gây ra yếu cơ) hay khiếm khuyết có thể rộng hơn, ảnh hưởng đến nhiều mô khác nhau như tim và các thận. Mức độ nghiêm trọng thay đổi từ tử vong ở trẻ nhỏ đến các rối loạn nhẹ mà không đe dọa tính mạng. Nhìn chung có 15 loại GSD đã được phát hiện; một vài trong số đó thì khá hiếm. Các loại phổ biến hơn của GSD thì được mô tả trong Bảng 1 và ba trong số các GSD phổ biến nhất được minh họa trong Hình 8.
VII. Tổng hợp bài viết

- Các tích trữ chính của glycogen trong cơ thể thì được tìm thấy trong cơ xương, nơi mà chúng đóng vai trò như là một nguồn nguyên liệu tích trữ cho sự tổng hợp ATP trong suốt quá trình co cơ và trong gan, nơi mà chúng được sử dụng để duy trì nồng độ glucose máu, đặc biệt là trong suốt giai đoạn đầu của sự nhịn ăn.
- Glycogen là một polymer phân nhánh mạnh của α-D-glucose.
- UDP-glucose, khối cấu trúc của glycogen, được tổng hợp từ glucose 1-phosphate và UTP bởi UDP-glucose pyrophosphorylase (Hình 14).
- Glucose từ UDP-glucose được chuyển đến các đầu tận không khử (nonreducing ends) của các chuỗi glycogen bởi glycogen synthase cần đoạn mồi, thành phần giúp hình thành nên các liên kết α(1→4). Đoạn mồi (primer) được tạo thành bởi glycogenin. Các nhánh được hình thành bởi amylo-α(1→4)→α(1→6)-transglycosylase (một 4:6 transferase), enzyme giúp chuyển một tập hợp 6 đến 8 gốc glucosyl từ đầu không khử của chuỗi glycogen (bẻ gãy một liên kết α[1→4]) và hình thành nên một liên kết α(1→6) với một gốc khác trong chuỗi.
- Glycogen phosphorylase phân tách các liên kết α(1→4) giữa các gốc glucosyl ở các đầu không khử của các chuỗi glycogen, tạo thành glucose 1-phosphate.
- Glucose 1-phosphate được chuyển thành glucose 6-phosphate bởi phosphoglucomutase.
- Trong cơ, glucose 6-phosphate đi vào trong quá trình đường phân. Trong gan, gốc phosphate được loại bỏ bởi glucose 6-phosphatase, giải phóng ra glucose tự do mà có thể được sử dụng để duy trì các mức glucose máu ở giai đoạn đầu của quá trình nhịn ăn.
- Một sự suy giảm phosphatase gây ra bệnh von Gierke và làm mất khả năng của gan trong việc cung cấp glucose tự do cho cơ thể trong suốt quá trình nhịn ăn. Nó tác động đến cả sự thoái hóa glycogen và quá trình tân tạo đường.
- Sự tổng hợp và thoái hóa glycogen thì được điều hòa qua lại để đáp ứng với các nhu cầu của toàn cơ thể bởi cùng các tín hiệu hormone, cụ thể là, mức insulin tăng thì tổng thể sẽ làm tăng quá trình tổng hợp glycogen và giảm quá trình thoái hóa glycogen, ngược lại, một sự tăng lên trong mức glucagon hay epinephrine sẽ gây ra các tác động trái ngược.
- Các enzyme quan trọng được phosphoryl hóa bởi các protein kinases, một số trong đó là phụ thuộc cAMP, một hợp chất tăng lên bởi glucagon và epinephrine. Các nhóm phosphate được loại bỏ bởi PP-1.
- Ngoài sự điều hòa cộng hóa trị này, glycogen synthase, phosphorylase kinase và phosphorylase thì cũng được điều hòa về mặt dị lập thể để đáp ứng với các nhu cầu của mô.
- Ở trạng thái ăn no, glycogen synthase được hoạt hóa bởi glucose 6-phosphate nhưng glycogen phosphorylase bị ức chế bởi glucose 6-phosphate cũng như là bởi ATP.
- Trong gan, glucose tự do cũng đóng vai trò như là một chất ức chế dị lập thể của glycogen phosphorylase.
- Sự tăng lên của canxi trong cơ trong suốt quá trình luyện tập thể thao và trong gan trong đáp ứng với epinephrine sẽ hoạt hóa phosphorylase kinase bởi sự liên kết với tiểu đơn vị CaM của enzyme. Điều này cho phép enzyme hoạt hóa glycogen phosphorylase, bằng cách đó, gây ra sự thoái hóa glycogen.
Các bạn có thể xem bài viết mới trên Facebook tại đây: https://www.facebook.com/profile.php?id=61550892771585
Các bạn có thể xem bài viết trước tại đây: https://docsachxyz.com/qua-trinh-tan-tao-duong-gluconeogenesis/
Cảm ơn các bạn đã theo dõi bài viết. Hẹn gặp lại các bạn trong các bài viết tiếp theo nhé !!!







{kind=link}